
|
Boletín de la Sociedad Geológica Mexicana Volumen 74, núm. 2, A230422, 2022 http://dx.doi.org/10.18268/BSGM2022v74n2a230422
|
 |
The paleoenvironmental significance of trace fossils from the Cárdenas Formation (Late Cretaceous, Maastrichtian) at Potrero del Carnero, San Luis Potosí, central Mexico
La importancia paleoambiental de las trazas fósiles de la Formación Cárdenas (Cretácico Tardío, Maastrichtiano) en Potrero del Carnero, San Luis Potosí, centro de México
Arturo Palma-Ramírez1, Roberto Carlos Maldonado-Sarabia2, Matthew R. Stimson3, Isabel López-Palomino4,*, Julio Reyes Romo-Ramírez5, Carlos Rene Muñóz-Jarillo6, Lisbeth Adenaly Badillo-Cadena5
1 Doctorado en Ciencias en Biodiversidad y Conservación, Área Académica de Biología. Universidad Autónoma del Estado de Hidalgo, Ciudad del Conocimiento, Carretera Pachuca-Tulancingo km 4.5, 42184, Pachuca, Hidalgo, Mexico.
2 Servicio Geológico Mexicano, Av. Quintana Roo 900, Col. República Oriente, 25280, Saltillo, Coahuila, Mexico.
3 Geology/ Paleontology Section, New Brunswick Museum, 277 Douglas Ave, Saint John New Brunswick E2K 1E5, Canada.
4 Léxico Estratigráfico de México, Departamento de Estratigrafía, Servicio Geológico Mexicano, Blvd. Felipe Ángeles km. 93.50-4, Col. Venta Prieta, 42083, Pachuca, Hidalgo, Mexico.
5 Gerencia Regional Centro, Servicio Geológico Mexicano, Av. Mariano Jiménez 465, Col. Alamitos, 78280, San Luis Potosí, San Luis Potosí, Mexico.
6 Maestría en Ingeniería Ambiental y Desarrollo Sustentable, Universidad Popular Autónoma de Puebla, 21 sur 1103, Barrio de Santiago, 72410, Puebla, Puebla, Mexico.
* Corresponding author: (I. López-Palomino) This email address is being protected from spambots. You need JavaScript enabled to view it.
How to cite this article:
Palma-Ramírez, A., Maldonado-Sarabia, R.C., Stimson, M.R., López-Palomino, I., Romo-Ramírez, J.R., Muñóz-Jarillo, C.R., Badillo-Cadena, L.A., 2022, The paleoenvironmental significance of trace fossils from the Cárdenas Formation (Late Cretaceous, Maastrichtian) at Potrero del Carnero, San Luis Potosí, central Mexico: Boletín de la Sociedad Geológica Mexicana, 74 (2), A230422. http://dx.doi.org/10.18268/BSGM2022v74n2a230422
Manuscript received: March 6, 2022; Corrected manuscript received: April 12, 2022; Manuscript accepted: April 25, 2022
ABSTRACT
This paper documents the occurrence of Skolithos linearis, Planolites beverleyensis, and Thalassinoides isp. in the Cárdenas Formation (Late Cretaceous), southeastern San Luis Potosí, central Mexico. This is the second record of S. linearis for this lithological unit, while P. beverleyensis and Thalassinoides isp. are documented for the very first time in this formation. The ichnoassemblage is referred to the Cruziana Ichnofacies which, in concert with associated invertebrate body fossils (Exogyra costata), suggest a shoreface setting with moderate- to low-energy conditions and episodic storms.
Keywords: Ichnofossils, Cruziana Ichnofacies, Late Cretaceous, Cárdenas Formation, San Luis Potosí, Mexico.
RESUMEN
El presente artículo documenta la presencia de Skolithos linearis, Planolites beverleyensis y Thalassinoides isp. en rocas de la Formación Cárdenas (Cretácico Tardío), sureste de San Luis Potosí, en el centro de México. La presencia de S. linearis representa el segundo registro de esta icnoespecie en esta unidad litológica, mientras que P. beverleyensis y Thalassinoides isp. son documentadas por primera vez en esta formación. El icnoensamble fue asignado a la Icnofacies de Cruziana, la cual, en conjunto con los invertebrados fósiles asociados (Exogyra costata), sugiere un ambiente de costa con condiciones de energía moderada a baja y episodios de tormenta.
Palabras clave: Icnofósiles, Icnofacies de Cruziana, Cretácico Tardío, Formación Cárdenas, San Luis Potosí, México.
- Introduction
The importance of invertebrate traces in paleontology, sedimentology, and stratigraphy has been demonstrated in several works (Pemberton et al., 2000, 2002; MacEachern et al., 2007, 2012). A remarkable characteristic of trace fossils is the amount of information that can provide concerning sedimentary environments (MacEachern et al., 2012). These biogenic structures give evidence of energy, sediment grain size, substrate type and consistency, food supply, water turbidity, oxygenation levels, and temperature (Allington-Jones et al., 2010; MacEachern et al., 2012).
Moreover, bioturbation intensity depends on various factors, such as trace maker population densities, types, rates, and depths of trace makers activities, as well as the period over which environmental conditions are favorable for colonization —the colonization window— (Rhoads, 1975; Pollard et al., 1993). When these colonization windows are only briefly open, individual trace fossils are more evident (Savdra, 2009).
The sensitivity of many animals to environmental conditions means that the trace fossils associations can be correspondingly more sensitive environmental indicators than inorganic sedimentary structures (Crimes, 1975). According to Allington-Jones et al. (2010), these associations tend to change progressively with depth, creating a basis for relative bathymetry.
The Cárdenas Formation has been studied for more than 100 years and several studies have demonstrated its paleontological significance documenting ammonites (Ifrim et al., 2005), brachiopods (Myers, 1968; Pérez-Martínez, 2010), corals (Navarro-Moctezuma, 2004; Baron-Szabo et al., 2006), crustaceans (Vega et al., 1995), echinoderms (Myers, 1968; Navarro-Moctezuma, 2004; Marín-Ávila, 2012), bivalves, gastropods (Böse, 1906), ostracods (Caus et al., 2002), rudists (Böse, 1906; Schafhauser et al., 2007; Oviedo-García, 2005; Pons et al., 2013), and foraminifers (Barker and Grimsdale, 1937; Carrillo-Bravo, 1971; Aguilar et al., 2002; Caus et al., 2002; Omaña et al., 2008, 2012, 2013). Its ichnological record includes traces of Ophiomorpha nodosa, Skolithos linearis, Diplocraterion parallelum, and Palaeophycus tubularis (Palma-Ramírez et al., 2019).
This paper documents for the very first time the trace fossils occurring in the Cárdenas Formation exposed at the Potrero del Carnero locality, southeastern San Luis Potosí, and discusses their paleoenvironmental implications.
- Geological setting and study area
Initially, the clastic rocks that constitute the Cárdenas Formation were first described by Böse (1906) in the surroundings of the city of Cárdenas, San Luis Potosí State, with the name of “División Cárdenas”. Later, Burckhardt (1930) used the term “Capas Cárdenas” for this sequence, and subsequently, Imlay (1944) elevated it to formation status. Myers (1968) divided this unit into three informal members (lower, middle, and upper) and proposed three biostratigraphic units based on its invertebrate body fossil assemblages (Durania ojanchalensis, Arcostrea aguilerae, and Tampsia floriformis Zones). Subsequent work by Carrillo-Bravo (1971) proposed four members (Member 1, Member 2, Member 3, and Member 4) according to the lithological features of this unit. The Cárdenas Formation is a 1,050 m thick unit, deposited during the Upper Cretaceous in a shallow marine setting characterized by fine siliciclastic rocks with interbedded limestone during a transgressive event, cropping out in an asymmetric syncline of the folded Sierra Madre Oriental (Myers, 1968; Carrillo-Bravo, 1971; López-Ramos, 1980).
This unit overlies the El Abra and Tamasopo formations (Myers, 1968; Santamaría-Orozco et al., 1990; Omaña et al., 2012), and it is unconformably overlain by the Tabaco Formation (Myers, 1968; López-Ramos, 1980; Schafhauser et al., 2007). In accordance with its stratigraphic relations and fossil content, the unit is Campanian-Maastrichtian in age (Carrillo-Bravo, 1971; Schafhauser et al., 2007; Omaña et al., 2012).
The study area is located in the Valles-San Luis Potosí Platform (VSLPP), a Mesozoic paleogeographic positive element where diverse evaporitic and marine Jurassic units, as well as shallow and reef Cretaceous carbonates, were deposited (Carrillo-Bravo, 1971). All these units underlay sandy and argillaceous with calcareous influence sediments (Carrillo-Bravo, 1971; Maldonado-Sarabia and Ríos-Vázquez, 2020). At Potrero del Carnero, Rayón municipality, southwestern San Luis Potosí State, the Cárdenas Formation conformably overlies the Tamasopo Formation (Figure 1; 21°52’18.84 N, 99°26’49.19 W).
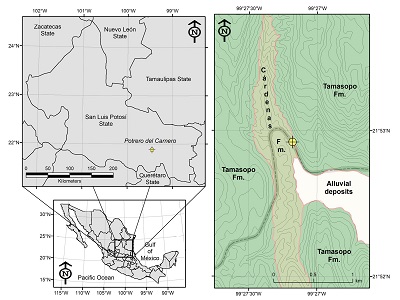 |
| Figure 1. Geographical location and geological map of the studied area. Modified from Maldonado-Sarabia and Ríos-Vázquez (2020). |
The only exposed outcrop is composed of a 13 m thick package which begins with limestone horizons of 0.50 to 1.10 m thick, light-brown on weathered surface and grey on fresh surface, with rudists, gastropods, and bivalve impressions. Also, there are sporadic thin horizons (around 0.1 m in thickness) of light-brown siltstone on fresh surfaces. Toward the top of the section, it consists of approximately 2.5 m of sandstone, gray in color on fresh surfaces and ocher on weathered surfaces, intercalated with gray lutite beds. Beds of sandstones range in thickness from 0.18 m to 0.55 m. The lutite is similar to the sandstones in color, with beds being 0.15 m to 0.36 m in thickness.
The trace fossils studied here come from the last 2.5 m of the section and are preserved in the sandstone and lutite beds. In the calcareous levels stratigraphically below the beds bearing the ichnofossils (Figure 2), Flores-Cadenas et al., (2018) recognized the benthic foraminifera Praechubbina, Chubbina, Cuneolina, as well as miliolids, and nezzazatids from the Campanian-Maastrichtian. However, the presence of the echinoderm Hemiaster and the ostreid Exogyra costata allow to correlate the ichnofossiliferous beds at Potrero del Carnero with the Arcostrea aguilerae biozone, in the middle portion of the Cárdenas Formation (early Maastrichtian) (Sohl and Kauffman, 1964; Myers, 1965, 1968; Vega et al., 1995).
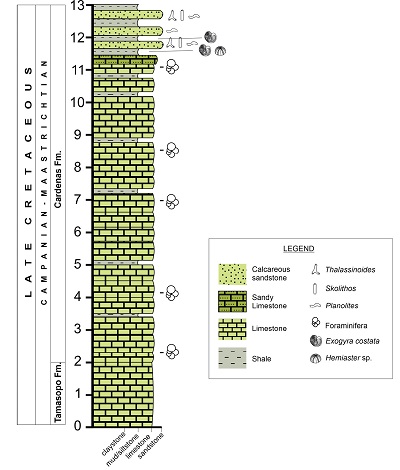 |
| Figure 2. Stratigraphic section of the Cárdenas Formation in Potrero del Carnero outcrop. Notice that trace fossils just crop near to the top of the studied section. |
- Methodology
The trace fossils come from a unique stratigraphic section measured and sampled at Potrero del Carnero locality (Figure 2). Since bed-by-bed sampling was not the main aim of the primary field survey, only a gross stratigraphic assignment is provided. Ichnofossils referred to and figured in this paper were photographed in the field and were not collected. In order to identify the ichnospecies we used the works published by Hadelman (1840), Billings (1862), Ehrenberg (1944), Buatois and Mángano (2011), and Fernández and Pazos (2012). Sedimentary structures, trace fossils, invertebrate fossils, and the intensity of bioturbation were considered for interpretation of the environment of deposition. We follow the scheme proposed by Taylor and Goldring (1993) to indicate the bioturbation intensity of the beds.
Bioturbation index (BI) refers to the determination of the extent of bioturbation or the degree to which the original physical sedimentary structures have been disrupted by biogenic reworking (Droser and Bottjer, 1989; Taylor and Goldring, 1993).
BI categorizes the extent of bioturbation into seven classes: BI=0, no bioturbation (0%); BI=1, sparse bioturbation with few discrete traces (1-4%); BI=2, low bioturbation in sediment that still has preserved sedimentary structures (5-30%); BI=3, moderate bioturbation and still distinguishable bedding boundaries (31-60%); BI=4, intense bioturbation, high trace-fossil density, common overlap of trace fossils, and primary sedimentary structures are mostly erased (61-90%); BI=5, sediment completely disturbed bedding and intense bioturbation (91-99%); BI=6, completely bioturbated and reworked sediment (100%).
- Systematic ichnology
Ichnogenus Planolites Nicholson, 1873
Planolites beverleyensis Billings, 1862
Description: Traces preserved as positive epirelief, straight to slightly curved, unbranched cylindrical burrows, horizontal to bedding planes. Burrows have smooth surface walls, are 0.8-2.3 cm diameter and up to 30 cm long. The passive fill in the burrow is constituted by gray lutite and the wall lining is not observed (Figures 3C to 3F).
 |
| Figure 3. Trace fossils of the Cárdenas Formation in Potrero del Carnero locality. A-B, Skolithos linearis; C, cross-section of Planolites beverleyensis; D-F, positive epirelief of Planolites beverleyensis; G-H, Thalassinoides isp.; I, positive epirelief of P. beverleyensis and Thalassinoides isp. |
Remarks: Planolites is ethologically interpreted as pascichnia of deposit feeders (Alpert, 1975; Fürsich, 1998; Knaust, 2010) produced by worms, insects, arthropods, mollusks, or infaunal holothuroids (Bromley, 1996; Uchman, 1998; Buatois and Mángano, 2002; Chen et al., 2011). Planolites is distinguished from Palaeophycus by the presence of wall linings and by a burrow-fill identical to the host rock (Pemberton and Frey, 1982; Mángano et al., 2005). Planolites beverleyensis can be separated from P. montanus by a minor diameter (P. montanus rarely exceeds 5 mm in diameter while P. beverleyensis rarely is less than 8 mm in diameter) and a markedly more sinuous and undulose of the latter (Pemberton and Frey, 1982).
Ichnogenus Skolithos Haldeman, 1840
Skolithos linearis Hadelman, 1840
Description: Straight, vertical to inclined, unbranched cylindrical burrows, with smooth walls. The burrow fill is homogeneous, structureless, medium to fine-grained sandstone, and is similar in composition to the host rock. Burrows are from 0.58 cm to 0.78 cm in width and have a maximum length of 6.238 cm; however, total length is difficult to determine because most specimens pass across the beds (Figure 3A and 3B).
Remarks: Skolithos is documented in a broad variety of environments but is most typical of shallow-water, high-energy settings (Pervesler et al., 2011). It is interpreted as dwelling (domichnia) and feeding (fodinichnia) burrows of annelids, sipunculids, phoronids and polychaete worms, crustaceans, anemones, and probably insects and spiders (Alpert, 1974; Bromley, 1996; Schrlif and Uchman, 2005). From a total of 35 recognized species of Skolithos, Alpert (1974) validated only five: S. verticalis (Hall), S. linearis Haldeman, S. magnus Howell, S. ingens Howell, and S. annulatus Howell. The main difference between S. verticalis with S. linearis and S. magnus is the greater diameter in the latter two cases (3–7 mm and 6–12 mm, respectively) (Alpert, 1974; Fernández and Pazos, 2012). On the other hand, S. ingens and S. annulatus, possess a greater diameter and exhibit characteristically ornamented walls (protuberances in S. ingens and annulation in S. annulatus), which are absent in S. verticalis (Fernández and Pazos, 2012).
Ichnogenus Thalassinoides Ehrenberg, 1944
Thalassinoides isp. Reit, 1932
Description: Horizontal, branched, cylindrical burrow system with dichotomous bifurcations, from 4 cm to 11 cm in length. Burrows have smooth surface walls, bifurcate in Y or T form with their diameters increasing at bifurcation points, giving the appearance of bulbs or irregular inflations. The tubular wall lining is not observed (Figure 3G to 3I).
Remarks: Thalassinoides burrows are considered as fodichnial, domichnial, and agrichnial structures (Bromley and Ekdale, 1984; Bromley, 1996; Miller, 2001; Ekdale and Bromley, 2003), attributed to decapod crustaceans, probably thalassinid shrimps or shrimp-like organism, crabs, sea anemones, and acorn worms (Bromley, 1996; Kim et al., 2002; Ekdale and Bromley, 2003; Pervesler and Uchmann, 2009; Chen et al., 2011, 2012). In accordance with Föllmi and Grimm (1990), the crustaceans producing Thalassinoides may survive transport in turbidity currents and build burrows under anoxic conditions during a limited number of days. Some of the features associated with Thalassinoides are shared with other burrows made by crustaceans namely Opiomorpha and Spongeliomorpha (Myrow, 1995). However, Thalassinoides can be distinguished from the others based on the differences among the burrows according to the specific behavioral patterns to specific taxa (Myrow, 1995). In particular, Thalassinoides lacks the pelletal lining of Ophiomorpha (Frey et al., 1978) and the scratchings of Spongeliomorpha (Bromley, 1967; Frey, 1970).
- Discussion
The trace fossil record from the Cárdenas Formation at Potrero del Carnero is characterized by burrows belonging to Planolites beverleyensis, Skolithos linearis, and Thalassinoides isp. The worms and crustaceans are the most probable trace makers of these traces. The aforementioned is congruent with that reported by Vega et al., (1995), who documented the presence of five decapod families, including two genera of callianassid shrimp. Individual traces are well preserved and are relatively abundant on some beds, which corresponds to BI=3 (moderate) for all bioturbated beds following the proposal of Taylor and Goldring (1993). This grade indicates a moderate level of bioturbation and still distinguishable bedding boundaries (Taylor and Goldring, 1993). The low diversity but high abundance of traces could be explained by stress factors such as oxygen depletion, brackish waters, hypersalinity, high energy, or biological influences such as predation (Taylor and Goldring, 1993; Buatois and Mángano, 2013).
Regarding the traces described herein, Skolithos is mainly recognized in shallow-water environments (Droser and Bottjer, 1989; Fillion and Pickerill 1990; Singh et al., 2008), but also rarely in non-marine settings and deep seas (Neto, 2007; Buatois and Mángano, 2011). This ichnogenus is also typical of the homonymous storm-related ichnofacies (Pemberton and Frey, 1984), and of high-energetic traction sedimentation (Mángano and Buatois, 2004). The ichnospecies Skolithos linearis is commonly interbedded between the lower offshore and the offshore transition facies, and it records a distinctive onshore-offshore trend (Mángano et al., 2005). Meanwhile Planolites is a eurybathic, extremely facies-crossing ichnogenus (Pemberton and Frey, 1982; Fillion and Pickerill, 1990). In particular, Planolites beverleyensis has been recorded in tempestites (Buatois and Mángano, 2011), as well as in lagoon and offshore bar settings (Tegan, 1992). Finally, Thalassinoides is a facies-crossing trace fossil, most typical of shallow-marine environment (Frey et al., 1978), associated with both firm and softgrounds (Myrow, 1995, MacEachern et al., 2007).
At Potrero del Carnero, the occurrence of vertical structures indicates opportunistic colonization events, whereas, the presence of horizontal structures is related to fair weather conditions (Perversler and Uchman, 2004; Benkhedda et al., 2021). On the one hand, the presence of Skolithos linearis could indicate a storm-related high-energy environment of the lower to middle shoreface (MacEachern et al., 2012), however, more detailed sedimentological studies are needed to reveal this setting. On the other hand, the presence of Thalassinoides could be interpreted as a shoreface depositional setting, which is supported by the presence of the ostreid Exogyra costata that finds favorable conditions in low energy environments (Myers, 1965). In addition, the co-occurrence of Thalassinoides and Planolites, which are essentially horizontal structures, suggest unconsolidated substrate experiencing relatively moderate to low energy in subtidal conditions (Malarkodi et al., 2009). Furthermore, simple morphologies, such as Thalassinoides, Skolithos, and Planolites, are related to salinity-stressed environments, dominating in brackish-water settings (Gingras et al., 2011).
This ichnological assemblage is dominated by dwelling (domichnia) and feeding (fodichnial) traces attributed to suspension- or deposit-feeding organisms, characteristics of the proximal Cruziana Ichnofacies (Fürsich, 1998; Buatois et al., 2002); which is transitional with the Skolithos Ichnofacies, and typical of the lower shoreface (Buatois and Mángano, 2011). Besides, the Cruziana Ichnofacies is associated with unconsolidated marine substrates, occurring mainly below fair-weather wave base and above storm weather wave base (Pemberton et al., 2001; Patel and Patel, 2015). Previously, based on an ichnological approach, Palma-Ramírez et al. (2019) document the Skolithos Ichnofacies in a section of the Cárdenas Formation located southwest of the studied area in this work. These authors suggest a marginal marine setting under low to high energy for early-late Maastrichtian rocks of the Cárdenas Formation exposed at Amoladeras, Rayón municipality, San Luis Potosí state. Due to the Cruziana Ichnofacies represents a deeper environment than Skolithos Ichnofacies, and because the age of the studied section here is older than the reported by the section studied by Palma-Ramírez et al., (2019), it is possible to infer that there was a decrease in the sea level throughout the early Maastrichtian in this area. Those above, agree with the documented transgressive-regressive conditions in the VSLPP, where sediment sources for the Cárdenas Formation were located to the west and northwest. The sediments filled an elongated shallow basin that was bordered to the east by a barrier that represented the beginning of folding and uplift of the Sierra Madre Oriental during the initial Laramide pulsations (Vega et al., 1995).
- Conclusions
The ichnological study of the Cárdenas Formation at Potrero del Carnero revealed a low ichnodiversity constituted by horizontal and sub-verticall burrows belonging to Skolithos linearis, Planolites beverleyensis, and Thalassinoides isp. Both, Planolites beverleyensis and Thalassinoides isp. represent the first record of these ichnospecies for this lithological unit. This ichnoassemblage corresponds to the proximal Cruziana Ichnofacies, suggesting moderate- to low-energy conditions in which food particles tend to accumulate on the sea floor rather than being kept in, with episodic storms.
Contributions of authors
Conceptualization: AP-R, IL-P, RCM-S; Data acquisition: AP-R, RCM-S, IL-P, JRR-R, CRM-J, LAB-C; Methodological development: AP-R, RCM-S, IL-P, JRR-R, CRM-J, LAB-C; Writing of the original manuscript: AP-R, RCM-S, IL-P; Graphic design: MS, RCM-S, IL-P; Fieldwork: AP-R, RCM-S, IL-P, JRR-R, CRM-J, LAB-C; Interpretation: AP-R, MS, RCM-S.
Financing
Financial support for this research came from the Mexican Geological Survey through the “Geological-Mining Cartography” Program and the “Mexican Stratigraphic Lexicon” Project. Matthew R. Stimson also acknowledges the support of the Natural Sciences and Engineering Research Council of Canada (NSERC) (funding reference number 547631) during the preparation of this and other manuscripts.
Acknowledgements
Thanks to Dr. Ana Laura Martínez-García (UAEH, Mexico), and Dr. Gabriela Pineda-Salgado (CONICET, Argentina) for their suggestions during the different stages of this work. The authors are grateful to Dr. Diana Elizabeth Fernandez (CONICET, Argentina) and one anonymous reviewer for their valuable advice and highlighted comments on the preliminary manuscript. Matthew R. Stimson thanks Nancy Jacque, David Wagner, Christopher Taylor, and Marion Homer for their moral and financial support during the writing of this and several other manuscripts.
Conflicts of interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
Aguilar, M., Bernaus, J.M., Caus, E., Hottinger, L., 2002, Lepidorbitoides minima Douvillé from Mexico, a foraminiferal index fossil for the Campanian: Journal of Foraminiferal Research, 32(2), 126-134. https://doi.org/10.2113/0320126
Allington-Jones, L., Braddy, S.J., Trueman, C.N., 2010, Palaeoenvironmental implications of the ichnology and geochemistry of the Westbury Formation (Rhaetian), Westbury-on-Severn, South-West England: Palaeontology, 53(3), 491-506. https://doi.org/10.1111/j.1475-4983.2010.00947.x
Alpert, S.P., 1974, Systematic review of the genus Skolithos: Journal of Paleontology, 48(4), 661-669.
Alpert, S.P., 1975, Planolites and Skolithos from the Upper Precambrian-Lower Cambrian White-Inyo Mountains, California: Journal of Paleontology, 49, 509-521.
Barker, R.W., Grimsdale, T.F., 1937, Studies of Mexican fossil foraminifera: The Annals and Magazine of Natural History, Ser. 10, 19(110), 161-178.
Baron-Szabo, R.C., Schafhauser, A., Götz, S., Stinnesbeck, W., 2006, Scleractinian corals from the Cárdenas Formation (Maastrichtian), San Luis Potosí, Mexico: Journal of Paleontology, 80(6), 1033-1046. https://doi.org/10.1666/0022-3360(2006)80[1033:SCFTCF]2.0.CO;2
Benkhedda, A., Defaflia, N., Cherif, A., Naimi, M.N., Mazouz, E.H., Palma-Ramírez, A., 2021, Shallow marine trace fossils from the Middle Miocene of the Tebessa Basin (NE Algeria) and their palaeonvironmental implications: Revista Brasileira de Paleontologia, 24(4), 323-335. https://doi.org/10.4072/rbp.2021.4.03
Billings, E., 1862, Palaeozoic Fossils. Volume 1: Montreal, Dawson Brothers, Geological Survey of Canada, 426p.
Böse, E., 1906, La Fauna de moluscos del Senomaniano de Cárdenas San Luis Potosí: Boletín del Instituto Geológico de México, 24, 91 p.
Bromley, R.G., 1967, Some observations on burrows of thalassinidean Crustacea in Chalk hardgrounds: Quarterly of Geological Society of London, 123, 157-182.
Bromley, R.G., 1996, Trace fossils. Biology, taphonomy and applications: London, Springer,361 p.
Bromley, R.G., Ekdale, A.A., 1984, Trace fossil preservation in flint in the European chalk: Journal of Paleontology, 58, 298-311.
Buatois, L.A., Mángano, M.G., Alissa, A., Carr, T.R., 2002, Sequence stratigraphic and sedimentologic significance of biogenic structures from a late Paleozoic marginal-to openmarine reservoir, Morrow Sandstone, subsurface of southwest Kansas, USA: Sedimentary Geology, 152, 99-132. https://doi.org/10.1016/S0037-0738(01)00287-1
Buatois, L.A., Mángano, M.G., 2011, Ichnology: The role of Organism-Substrate Interactions in Space and Time: Cambridge, Cambridge University Press, 358p.
Buatois, L.A., Mángano, G., 2013, Ichnodiversity and ichnodisparity: significance and caveats: Letahia, 46, 281-292. https://doi.org/10.1017/CBO9780511975622
Burckhardt, C., 1930, Etude Synthetíque sur le Mésozoïque Mexicain: Memoires de la Société Paléontologique Suisse, Vol. II. 231-233.
Carrillo-Bravo, J., 1971, La Plataforma Valles-San Luis Potosí. Ciudad de México: Boletín de la Asociación Mexicana de Geólogos Petroleros, 23(1-6), 106p.
Caus, E., Tambareau, Y., Colin, J.P., Aguilar, M., Bernaus, J.M., Gómez-Garrido, A., Brusset, S., 2002, Upper Cretaceous microfauna of the Cárdenas Formation, San Luis Potosí, NE Mexico. Biostratigraphical, palaeoecological, and palaeogeographical significance: Revista Mexicana de Ciencias Geológicas, 19(2), 137-144.
Chen, Z.Q., Tong, J., Fraiser, M.L., 2011, Trace fossil evidence for restoration of marine ecosystems following the end-Permian mass extinction in the Lower Yangtze region, South China: Palaeogeography, Palaeoclimatology, Palaeoecology, 299, 449-474. https://doi.org/10.1016/j.palaeo.2010.11.023
Chen, Z.-Q., Fraiser, M.L., Bolton, C., 2012, Early Triassic trace fossils from Gondwana Interior Sea: implications for ecosystem recovery following the end-Permian mass extinction in south high-latitude region: Gondwana Research, 22, 238-255. https://doi.org/10.1016/j.gr.2011.08.015
Crimes, R.P., 1975, The production and preservation of trilobite resting and furrowing traces: Lethaia, 8,35-48.
Droser, M.L., Bottjer, D.J., 1989, Ichnofabric of sand-stones deposited in high-energy nearshore environment: measurement and utilization: Palaios, 4, 598-604.
Ehrenberg, K., 1944, Ergianzende Bemerkungen zu den seinerzeit aus dem Miozan von Burgschleintz beschriebenen Gangkeren und Bauten dekapoder Krebse: Palaeontologische Zietschrift, 23, 354-359.
Ekdale, A.A., Bromley, R.G., 2003, Paleoethologic interpretation of complex Thalassinoides in shallow-marine limestones, Lower Ordovician, southern Sweden: Palaeogeography, Palaeoclimatology, Palaeoecology, 192, 221-227. https://doi.org/10.1016/S0031-0182(02)00686-7
Fernández, D.E., Pazos, P.J., 2012, Ichnology of marginal marine facies of the Agrio Formation (Lower Cretaceous, Neuquén Basin, Argentina) at its type locality: Ameghiniana, 49, 505-524. https://doi.org/10.5710/AMGH.23.7.2012.439
Fillion, D., Pickerill, R.K., 1990, Ichnology of the Upper Cambrian? to Lower Ordovician Bell Island and Wabana groups of eastern Newfoundland, Canada: Palaeontographica Canadiana, 7, 1-119.
Föllmi, K.B., Grimm, K.A., 1990, Doomed pioneers: Gravity flow deposition and bioturbation in marine oxygen deficient environments: Geology, 18, 1069-1072.
Flores-Cadenas, B.E., Monier-Castillo, A., López-Palomino, I., Palma-Ramírez, A., Pérez-Martínez, M., 2018, Informe sobre el estudio micropaleontológico de muestras (láminas delgadas), correspondientes a la localidad El Carnero, Carta Geológico-Minera Tamasopo (F14-C18), escala 1:50,000, O.T. 14965: Pachuca, Hidalgo, Servicio Geológico Mexicano, Departamento de Estratigrafía, internal report, 9 p.
Frey, R.W., 1970, Trace fossils of Fort Hays Limestone Member of Niobrara Chalk (Upper Cretaceous), west-central Kansas: Paleontology Contributions of the University of Kansas, 53, 41 p.
Frey, R.W., Howard, J.D., Pryor, W.A., 1978, Ophiomorpha: Its morphological, taxonomic, and environmental significance: Palaeogeography, Palaeoclimatology, Palaeoecology, 23,511- 528. https://doi.org/10.1016/0031-0182(78)90094-9
Fürsich, F.T., 1998, Environmental distribution of trace fossils in the Jurassic of Kachchh (Western India): Facies, 39, 243-272. https://doi.org/10.1007/BF02537019
Gingras, M.K., MacEachern, J.A., Dashtgard, S.E., 2011, Process ichnology and the elucidation of physico-chemical stress: Sedimentary Geology, 237, 115-134. https://doi.org/10.1016/j.sedgeo.2011.02.006
Haldeman, S.S., 1840, A monograph of the Limniades, or freshwater univalve shells of North America, Supplement to number 1: Philadelphia, J. Dobson, 3 p.
Ifrim, C., Stinnesbeck, W., Schafhauser, A., 2005, Maastrichtian shallow-water ammonites of northeastern Mexico: Revista Mexicana de Ciencias Geológicas, 22(1), 48-64.
Imlay, R.W., 1944, Cretaceous Formation of Central America and México: Bulletin of the American Association of Petroleum Geologists, 28(8), 1077-1195. https://doi.org/10.1306/3D9336A8-16B1-11D7-8645000102C1865D
Kim, J.Y., Kim, K.S, Pickerill, R.K., 2002, Cretaceous non-marine trace fossils from the Hasandong and Jinju Formations of the Namhae Area, Kyongsangnamdo, southeast Korea: Ichnos, 9, 41-60. https://doi.org/10.1080/10420940216414
Knaust, D., 2010, The end-Permian mass extinction and its aftermath on an equatorial carbonate platform; insights from ichnology: Terra Nova, 22, 195-202. https://doi.org/10.1111/j.1365-3121.2010.00934.x
López-Ramos, E., 1980, Geología de México. Volume II: México, D.F., Trillas, 454p.
MacEachern, J.A., Pemberton, S.G., Gingras, M.K., Bann, K.L., 2007, The Ichnofacies Paradigm: A Fifty-Year Retrospective, in Miller III, W. (ed.), Trace fossils: concepts, problems, prospects: New York, Elsevier, 52-77. https://doi.org/10.1016/B978-044452949-7/50130-3
MacEachern, J.A., Bann, K.L., Gingras, M.K., Zonneveld, J.P., Dashtgard, S.E., Pemberton, S.G., 2012, The ichnofacies paradigm, in Knaust, D. . Bromley, R.G. (eds.), Trace fossils as indicators of sedimentary environments, (Developments in Sedimentology 64): Oxford, Elsevier, 103–138. https://doi.org/10.1016/B978-0-444-53813-0.00004-6
Malarkodi, N., Patel, S.J., Fayazudeen, P.J., Mallikarjuna, U.B., 2009, Palaeoenvironmental significance of trace fossils from the Paleocene sediments of the Pondicherry area, South India: Journal Geological Society of India, 74, 738-748. https://doi.org/10.1007/s12594-009-0190-x
Maldonado-Sarabia, R.C., Ríos-Vázquez, J.R., 2020, Informe de la Carta Geológico-minera Tamasopo F14-C18, escala 1:50,000, estado de San Luis Potosí: Pachuca, Hidalgo, Mexico, Servicio Geológico Mexicano, technical report, 77 p.
Mángano, M.G., Buatois, L.A., 2004, Ichnology of Carboniferous tide-influenced environments and tidal flat variability in the North American Midcontinent, in McIlroy, D. (eds.), The application of ichnology to palaeoenvironmental and stratigraphic analysis: Geological Society of London, Special Publications, 228(1), 157-178. https://doi.org/10.1144/GSL.SP.2004.228.01.09.
Mángano, M.G., Buatois, L.A., Muniz-Guinea, F., 2005, Ichnology of the Alfarcito Member (Santa Rosita Formation) of northwestern Argentina: animal-substrate interactions in a lower Paleozoic wave-dominated shallow sea: Ameghiniana 42, 641-668.
Marín-Ávila, A.A., 2012, Taxonomía de los equinodermos de dos localidades (La Calzada y Puerto de Sta. Gertrudis) de la Formación Cárdenas en el municipio de Cd. del Maíz, S.L.P.: San Luis Potosí, México, Universidad Autónoma de San Luis Potosí, Bachelor’s thesis, 52p.
Miller W., III, 2001, Thalassinoides-Phycodes compound burrow systems in Paleocene deep-water limestone, Southern Alps of Italy: Palaeogeography, Palaeoclimatology, Palaeocology, 170, 149-156. https://doi.org/10.1016/S0031-0182(01)00234-6
Myers, R.L., 1965, Biostratigraphy of the Cárdenas Formation (Upper Cretaceous) San Luis Potosí, Mexico: Texas, USA, The University of Texas, PhD dissertation, 71p.
Myers, R.L., 1968, Biostratigraphy of the Cárdenas Formation (Upper Cretaceous) San Luis Potosí, Mexico: Paleontología Mexicana 24, 1-89.
Myrow, P.M., 1995, Thalassinoides and the enigma of Early Paleozoic open-framework burrow systems: Palaios, 10, 58-74. https://doi.org/10.2307/3515007
Navarro-Moctezuma, A., 2004, Descripción del Phyllum Celenterata y Echinodermata del miembro superior de la Formación Cárdenas del Cretácico Tardío de la localidad tipo, Cárdenas, S.L.P.: San Luis Potosí, México, Universidad Autónoma de San Luis Potosí, Bachelor’s thesis, 112p.
Netto, R.G., 2007, Skolithos-dominated piperock in non-marine environments. An example from the Triassic Caturrita Formation, Southern Brazil, in Bromley, R.G., Buatois, L.A., Mángano, G., Genise, J.F., Melchor, R.N. (eds), Sediment-organisms interactions: A multifaceted ichnology: SEPM Special Publication, 88, 109-121. https://doi.org/10.2110/pec.07.88.0107
Nicholson, H.A., 1873, Contributions to the study of the errant annelides of the Older Palaeozoic rocks: Proceedings of the Royal Society of London, 21, 288-290.
Omaña, L., Pons, J.M., Alencáster, G., 2008, Latest Cretaceous foraminifera from the Cárdenas Formation, San Luis Potosí, Mexico: Biostratigraphical, paleoenvironmental and paleobiogeographical significance: Micropaleontology 54, 445–462. https://doi.org/10.47894/mpal.54.5.04
Omaña, L., Alencáster, G., Torres-Hérnandez, J.R., López-Doncel, R., 2012, Morphological abnormalities and dwarfism in Maastrichtian foraminifers from the Cárdenas Formation, Valles–San Luis Potosí Platform, Mexico: evidence of paleoenvironmental stress: Boletín de la Sociedad Geológica Mexicana, 64(3), 305-318. https://doi.org/10.18268/BSGM2012v64n3a4
Omaña, L., López-Doncel, R., Torres-Hernández, J.R., Alencáster, G., 2013, Biostratigraphy and paleoenvironmental of the Cenomanian/Turonian boundary interval based on foraminifera from the western part of the Valles-San Luis Potosí Platform, Mexico: Micropaleontology, 58(6), 457-485. https://doi.org/10.47894/mpal.58.6.01
Oviedo-García, A., 2005, Rudistas del Cretácico Superior del centro al sureste de México: Spain, Universitat Autònoma de Barcelona, Ph.D. dissertation, 123p.
Palma-Ramírez, A., Maldonado-Sarabia, R.C., Stimson, M., 2019, Marginal marine trace fossils from the Cárdenas Formation (Maastrichtian), Rayón municipality, San Luis Potosí, Central Mexico: Revista Brasileira de Paleontologia, 22(2), 89-96. https://doi.org/10.4072/rbp.2019.2.01
Patel, S.J., Patel, N.J., 2015, Sedimentological and palaeoecological significance of the trace fossils of the Jurassic rocks of the Jhura Dome, Mainland Kachchh, western India: Volumina Jurassica, 13(2), 101-140. https://doi.org/10.5604/17313708.1185696
Pemberton, S.G., Frey, R.W., 1982, Trace fossil nomenclature and the Planolites-Palaeophycus dilemma: Journal of Paleontology, 56(4), 843-881.
Pemberton, S.G., Frey, R.W., 1984, Ichnology of the storm-influenced shallow marine sequence: Cardium Formation (Upper Cretaceous) at Seebe, Alberta, in Stoot, D.F., Glass, D.J. (eds.), The Mesozoic of Middle North America: Canadian Society of Petroleum Geologists Memoirs, Alberta, 9, 281-304.
Pemberton, S.G., MacEachern, J.A., Gingras, M.K., Zang, J., 2000, Significance of ichnofossils to genetic stratigraphy: examples from the Cretaceous of Alberta, Canada. Science in China, Series D, Earth Sciences, 43,541–560. https://doi.org/10.1007/BF02875316.
Pemberton, S.G., Spila, M., Pulham, A.J., Saunders, T., MacEachern, J.A., Robbins, D., Sinclair, I.K., 2002, Ichnology and sedimentology of shallow to marginal marine systems. New found land, The Geological Association of Canada, 343 p.
Pemberton, S.G., Spila, M., Pulham, A.J., Saunders, T., MacEachern, J.A., Robbins, D., Sinclair I.K., 2001, Ichnology and sedimentology of shallow to marginal marine systems: Ontario, Geological Association of Canada. https://doi.org/10.1669/0883-1351(2003)018%3C0471:IASOST%3E2.0.CO;2
Pérez-Martínez, M.B., 2010, Identificación de 4 especies de brachiopodos de la Formación Cárdenas, en la localidad La Calzada, municipio de Ciudad del Maíz, como primer reporte de este grupo: San Luis Potosí, Mexico, Universidad Autónoma de San Luis Potosí, Bachelor’s thesis, 45p.
Pervesler, P., Uchman, A., 2009, A new Y-shaped trace fossil attributed to upogebiid crustaceans from Early Pleistocene of Italy: Acta Palaeontologica Polonica, 54, 135-142. https://doi.org/10.4202/app.2009.0114
Pervsersler, P., Roetzel, R., Uchman, A., 2011, Ichnology of shallow sublittoral siliciclastics of the Burgschleinitz Formation (Lower Miocene, Eggenburgian) in the Alpine-Carpathian Foredeep (NE Austria): Austrian Journal of Earth Sciences, 104(1), 81-96.
Pollard, J.E., Goldring, R., Buck, S.G., 1993, Ichnofabrics containing Ophiomorpha: significance in shallow-water facies interpretation: Journal of the Geological Society, 150,149–164. https://doi.org/10.1144/gsjgs.150.1.0149
Pons, J.M., Vicens, E., Oviedo, A., Aguilar, J., García-Barrera, P., Alencáster, G., 2013, The rudist fauna of the Cárdenas Formation, Maastrichtian, San Luis Potosí State, Mexico: Journal of Paleontology, 84(4), 726-754. https://doi.org/10.1666/12-116
Rhoads, D.C., 1975, The paleoecological and environmental significance of trace fossils, in Frey, R.W. (ed.), The study of trace fossils. A synthesis of principles, problems and procedures in ichnology: Berlin, Springer Verlag, 147-160. https://doi.org/10.1007/978-3-642-65923-2_9
Santamaría-Orozco, D., Arenas-Partida, R., Escamilla-Herrera, A., 1990, Normalización de la nomenclatura estratigráfica en las cuencas mesozoicas de México (Etapa I: Zona Norte. Cretácico): México, D.F., Instituto Mexicano del Petróleo, 140 p.
Savdra, C.E., 2009, Taphonomy of trace fossils, in Miller III, W. (ed.), Trace Fossils. Concepts, Problems, Prospects: Amsterdam, Elsevier, 92-109.
Schafhauser, A., Götz, S., Stinnesbeck, W., 2007, Rudist decline in the Maastrichtian Cardenas Formation (East-central Mexico): Palaeogeography, Palaeoclimatology, Palaeoecology, 251, 210-221. https://doi.org/10.1016/j.palaeo.2007.03.047
Schlirf, M., Uchman, A., 2005, Revision of the ichnogenus Sabellarifex Richter, 1921 and its relationship to Skolithos Haldeman, 1840 and Polykladichnus Fursich, 1981: Journal of Systematic Palaeontology, 3, 115-131. https://doi.org/10.1017/S1477201905001550
Singh, R.H., Rodríguez-Tovar, F.J., Ibotombi, S., 2008, Trace fossils of the Upper Eocene-Lower Oligocene transition of the Manipur, Indo-Myanmar Ranges (North-east India): Turkish Journal of Earth Sciences, 17, 821-834.
Sohl, N.F., Kauffman, E.G., 1964, Giant Upper Cretaceous oysters from the Gulf Coast and Caribbean: Geological Survey Professional Paper 483-H, 22p.
Taylor, A.M., Goldring, R., 1993, Description and analysis of bioturbation and ichnofabric: Journal of the Geological Society, 150, 141-148.
Tegan, J.R., 1992, Ichnology and paleoenvironments of the Lowville, Napanee, and lower Kings Falls Limestone (Upper Ordovician), Ingha, Mills, Mohawk Valley, New York, in 5th Keck Symposium: Virginia, USA, KECK Geology Consortium, 203-208.
Uchman, A., 1998, Taxonomy and ethology of flysch trace fossils: revision of the Marian Książkiewicz Collection and studies of complementary material: Annales Societatis Geologorum Poloniae, 68, 105-218.
Vega, F.J., Feldmann, R.M., Sour-Tovar, F., 1995, Fossil crabs (Crustacea: Decapoda) from the Late Cretaceous Cárdenas Formation, east-central Mexico: Journal of Paleontology, 69(2), 340-350. https://doi.org/10.1017/S0022336000034661