
|
Boletín de la Sociedad Geológica Mexicana Volumen 72, núm. 1, 2020, p. 1-19 |
 |
Nuevos registros de mamíferos pleistocénicos de San Mateo Huexoyucán, Tlaxcala y el segundo registro de juveniles de Mammuthus en México.
New records of Pleistocene mammals from San Mateo Huexoyucán, Tlaxcala and the second Mammuthus juvenile registry in Mexico
Miguel Sánchez-Salinas1, Eduardo Jiménez-Hidalgo2, J. Alberto Cruz1, Carlos Castañeda-Posadas1,*
1 Benemérita Universidad Autónoma de Puebla, Facultad de Ciencias Biológicas, Laboratorio de Paleontología, Escuela de Biología, Blvd. Valsequillo y Av. San Claudio, Edificio 112-A, Ciudad Universitaria, Jardines de San Manuel, 72570, Puebla, México.
2 Laboratorio de Paleobiología, Instituto de Recursos, Universidad del Mar, campus Puerto Escondido, Laboratorio de Paleobiología, Km 2.5, carretera Puerto Escondido-Sola de Vega, 71980, Puerto Escondido, Oaxaca, México.
*Autor para correspondencia: (C. Castañeda-Posadas)
This email address is being protected from spambots. You need JavaScript enabled to view it.
Cómo citar este artículo: Sánchez-Salinas, M., Jiménez-Hidalgo, E., Cruz, J.A., Castañeda-Posadas, C., 2020, Nuevos registros de mamíferos pleistocénicos de San Mateo Huexoyucán, Tlaxcala y el segundo registro de juveniles de Mammuthus en México: Boletín de la Sociedad Geológica Mexicana, 72 (1), A240919. http://dx.doi.org/10.18268/BSGM2020v72n1a240919
Resumen
En este trabajo se describen formalmente fósiles provenientes de la zona de San Mateo Huexoyucán-Tlaxcala, México. Los especímenes fueron colectados desde 2011 e incluyen elementos craneales y postcraneales de Megafauna pleistocénica. La identificación de los elementos fósiles nos permite postular que corresponden a siete especies previamente reportadas para la zona de estudio (Bison antiquus, Camelops hesternus, Cuvieronius hyodon, Equus conversidens, Equus mexicanus y Mammuthus sp.), destaca la presencia de material fósil que corresponde a ejemplares juveniles de proboscideos y gliptodontes (cf. Glyptotherium floridanum).
Palabras clave: Megafauna, Rancholabreano, Tlaxcala, Centro de México, Pleistoceno.
Abstract
The fossils from San Mateo Huexoyucan, Tlaxcala, Mexico are formally described in this work. The specimens collected since 2011 include cranial and postcranial elements of Pleistocene Megafauna. The identification of the elements allows to postulate that they correspond to six previously reported species for the study area (Bison antiquus, Camelops hesternus, Cuvieronius hyodon, Equus conversidens, E. mexicanus and Mammuthus sp.), highlighting the presence of fossil material that corresponds to a juvenile individual of proboscideans (Mammuthus sp.) and glyptodonts (cf. Glyptotherium floridanum).
Keywords: Megafauna, Rancholabrean, Tlaxcala, Central Mexico, Pleistocene.
- Introducción
La edad de mamíferos de Norteamérica conocida como Rancholabreano, que corresponde al Pleistoceno, se define como el intervalo entre los 210000‒160000 a 9500 años A.P., la cual abarca una distribución geográfica desde los 55° de latitud norte hacia el sur de Norteamérica.
Biocronológicamente, inicia con la aparición en el registro fósil del género Bison y finaliza con la extinción de la Megafauna pleistocénica (Bell et al., 2004). En zoología terrestre, el término Megafauna (del griego megas=grande y del latín fauna=animal) se refiere a animales “gigantes”, “muy grandes” o “grandes” que superan los 44 kg (Stuart, 1991); este término es asociado con la fauna de los periodos Paleógeno, Neógeno y Cuaternario. Los animales grandes o gigantes que se consideran arquetípicos de la última era de hielo como los mamuts en el hemisferio norte y los gliptodontes en América del Sur, forman parte de esta Megafauna (Fariña et al., 2013). El estudio de los megamamíferos del Pleistoceno es importante para la interpretación de los procesos evolutivos que tuvieron lugar en el pasado, así como para el estudio de la relación de estos seres con el ambiente (Donovan y Paul, 1998; Gómez-Pérez y Carbot-Chanona, 2012).
La Megafauna de México está bien estudiada a nivel país (Arroyo-Cabrales et al., 2008; Ceballos et al., 2010; Ferrusquía-Villafranca et al., 2010, 2017); sin embargo, existen estados como Tlaxcala donde estos estudios son escasos, por lo que en este trabajo se revisará el material depositado y no publicado de la Colección Paleontológica de la Benemérita Universidad Autónoma de Puebla (BUAPAL), para aumentar el conocimiento de la diversidad pleistocénica del estado de Tlaxcala.
- Antecedentes
Para Tlaxcala se tienen registros sobre asociación faunística pleistocénica provenientes de San Mateo Huexoyucán (Sánchez-Salinas et al., 2016), estudios de gliptodontes (Ramírez-Cruz y Montellano-Ballesteros, 2014) y équidos pleistocénicos (Priego-Vargas et al., 2017). Bonilla-Toscano, en su tesis de licenciatura (2011), describió ejemplares rescatados en la barranca de San Mateo Huexoyucán, reportando cuatro taxa: Equus, Bison, Mammuthus y Camelidae. En el presente trabajo se revisó parte de este material, además se incluyeron nuevos materiales colectados durante la temporada de excavación 2016-2017.
- Área de estudio
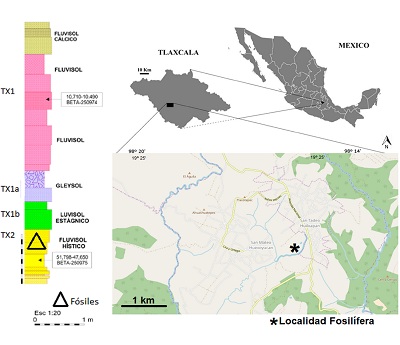
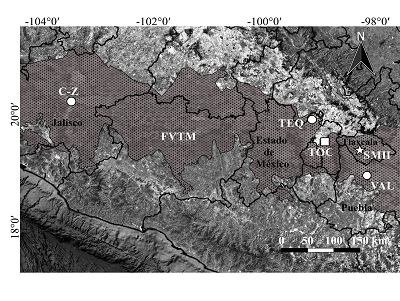
La comunidad de San Mateo Huexoyucán se ubica al norte del municipio de Panotla, en las carreteras federales en dirección San Simeón Xipetzingo, en el estado de Tlaxcala (INAFED, 2010). El pedocomplejo de la Barranca de San Mateo Huexoyucán se ha clasificado como un fluvisol hístico; sus niveles fosilíferos forman parte de una secuencia de fluvisoles (Figura 1) con una datación estimada de radiocarbono de 46320 ± 870 años A.P. y una edad calibrada de 51798 ‒ 47650 años A.P., correspondiente al Rancholabreano en el Pleistoceno tardío (Sánchez-Salinas et al., 2016).
- Material y métodos
Se revisó el material depositado y no publicado de la Colección Paleontológica de la Benemérita Universidad Autónoma de Puebla (BUAPAL). Los ejemplares fueron identificados por medio de anatomía comparada y utilizando bibliografía especializada en mamíferos pleistocénicos (Oslen, 1960; Webb, 1965; Gillette y Ray, 1981; Lucas et al., 2011 y Fisher et al., 2014). Se identificaron cuatro órdenes (Artiodactyla, Proboscidea, Perissodactyla y Xenathra) y seis familias (Bovidae, Camelidae, Elephantidae, Equidae, Glyptodontidae y Gomphotheriidae), las cuales se describen a continuación.
|
Figura 1. Mapa y columna estratigráfica del área de estudio (modificado de Sánchez-Salinas et al., 2016). |
- Resultados
Orden ARTIODACTYLA Owen, 1848
Familia BOVIDAE Gray, 1821
Género Bison Smith, 1827
Bison antiquus Leidy, 1852
Figura 2
Material referido. BUAPALZ-979, dentario izquierdo.
Descripción. El elemento es largo y esbelto, de 37.7 cm de largo, sin incisivos, con un foramen mental profundo y ovalado de 1.4 cm de diámetro, el diastema es esbelto y no está elevado (Figura 2). La serie post-canina está completa: el p2 es triangular y con una fosetida en forma de “V”; la corona del p3 está fragmentada y tiene una fosetida en forma de “U” y otra de “V”, mientras que en el p4 sus fosetidas tienen en forma de “U”, “V” y otra ovoide, con costillas bien definidas; los molares están desgastados, robustos y tienen el borde labial en forma de “M”; el m1 está fragmentado, las fosetidas tienen forma de “8”, el ectostilido tiene forma ovoide; el m2 tiene fosetas en forma de “8”, con ectostilido roto y con forma de gota. El grado de desgaste m3 sugiere un individuo adulto, las fosetidas tienen forma de “8” y el ectostilido es triangular y desgastado.
|
Figura 2. BUAPALZ-979, dentario izquierdo de Bison antiquus. La escala representa 5 cm. Abreviaturas: DIA, diastema; FOM, foramen mental, MMA, muesca mandibular; PC, proceso coronoideo; SIN, sínfisis; m, molar inferior; p, premolar inferior. |

La rama dental mide 19.3 cm de longitud; el proceso coronoideo es curveado y esbelto, la muesca mandibular es profunda y en forma de gota, mientras que el proceso condilar de la mandíbula está fragmentado y cóncavo. En la vista lingual de la mandíbula se puede apreciar el foramen mandibular fragmentado y profundo, seguido por una pequeña depresión convexa. También conserva parte de la sínfisis, la cual tiene varios alveolos en la superficie.
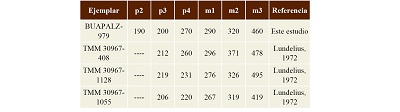
Discusión. El ejemplar BUAPALZ-979 se identificó como un bóvido ya que en Bison y Bos la articulación mandibular es cóncava y plana, el proceso coronoideo es robusto, terminando en una punta; los molares son hispsodontos y se pliegan en forma de “M” y “V”; las fosetas de los molares tienen forma de 8 (Schmid, 1972; Hillson, 2005; France, 2009). A diferencia de Equus la articulación mandibular es convexa y el proceso coronoideo es redondo (Schmid, 1972; France, 2009). En ejemplares adultos el m3 presenta un pliegue del exoestilo triangular con forma de gota (Schmid, 1972; Hillson, 2005) tal como se puede apreciar en el ejemplar BUAPALZ-979. El diastema mandibular en Bos es ancho y elevado, pero no afilado, mientras que en Bison el diastema tiene un margen afilado y un borde ligeramente convexo (Schmid, 1972; Jiménez-Hidalgo et al., 2013); estas características identifican al ejemplar BUAPALZ-979 perteneciente al género Bison. Los molares inferiores se compararon con ejemplares de mandíbulas de Bison antiquus de Ingleside, Texas, reportados por Lundelius (1972), y se encontró que los molares del ejemplar estudiado son similares en tamaño a los de Bison antiquus (Tabla 1), por lo que el ejemplar BUAPALZ-979 se identificó como perteneciente a esta especie de bisonte.
|
Tabla 1. Medidas en mm de los molares inferiores del Ejemplar BUAPALZ-979 y otros dentarios de Bison antiquus reportados por Lundelius, 1972.
|
Familia CAMELIDAE Gray, 1821
Camelops Leidy, 1854
Camelops hesternus Leidy, 1873
Figura 3
Material referido. BUAPALZ-983, fragmento de cráneo.
Descripción. Cráneo fragmentado largo y grande de 15 cm de longitud y 8.3 cm de ancho, que conserva parte del hueso escamoso, el hueso occipital y la cresta supraoccipital.
En vista dorsal, el escamoso es cóncavo y tiene dos forámenes, uno pequeño y ovalado, mientras que el otro es grande, profundo y cercano al meato auditivo externo; se puede ver la cresta sagital externa la cual es gruesa; la cresta occipital está fragmentada. En vista lateral se puede observar un foramen accesorio del meato temporal, el cual es grande y profundo, seguido por el meato auditivo externo de 2.9 cm, amplio, plano y profundo, rodeado por un labio prominente y fragmentado; detrás de éste se encuentra el foramen mastoideo reniforme. En la parte posterior el ejemplar BUAPALZ-983 conserva un cóndilo occipital plano y triangular de 3.5 cm; el proceso paraoccipital tiene forma de gancho, es plano lateralmente y sobrepasa el cóndilo occipital largo y triangular (Figura 3).
Discusión. El ejemplar BUAPALZ-983 comparte características de camélidos: la cresta paraoccipital del ejemplar de Tlaxcala es amplia como en la llama, el foramen mastoideo reniforme y la apófisis paramastoidea es más pronunciada que en Equus (Galotta y Galotta, 1988). La morfología del cráneo de Camelops es más cercana al cráneo de la llama sudamericana Lama que al camello bactriano euroasiático Camelus (Webb, 1965). En Camelops hesternus, el meato auditorio externo es largo, plano anteroposteriomente y rodeado por un labio prominente; también presenta dos forámenes subescamosos, el primero es pequeño y el segundo más grande y tiende a ser ovalado; la parte dorsal del occipital de Camelops es extensa, la cresta se proyecta posteriormente sobre los cóndilos y la fosa mastoidea es considerablemente profunda, el proceso paraoccipital es en forma de gancho anteromedialmente y aplanado lateralmente, los cóndilos occipitales son alargados y cóncavos (Webb, 1965). El ejemplar BUAPALZ-983 comparte estos caracteres que lo identifican como Camelops hesternus.
|
Figura 3. BUAPALZ-983, fragmento de cráneo de Camelops hesternus. A) vista craneal; B) vista lateral. La escala representa 5cm. Abreviaturas: COC, cóndilo occipital; CRO, cresta occipital; ESC, hueso escamoso; FO, foramen accesorio; MA, meato auditivo; OCC, occipital, PAO, proceso paraoccipital. |

Orden PROBOSCIDEA Illiger, 1811
Familia GOMPHOTHERIIDAE Cabrera, 1929
Género Cuvieronius Osborn, 1923
Cuvieronius hyodon Fischer, 1814
Figura 4
Material referido. BUAPALZ-972, calcáneo fragmentado.
Descripción. Calcáneo de 16 cm de largo y 21.5 cm de alto, carece de sustentáculo y solo conserva la tuberosidad calcánea, la cual es alargada y redondeada, con cuello corto y presenta la faceta para el astrágalo elíptica y fragmentada (Figura 4a).
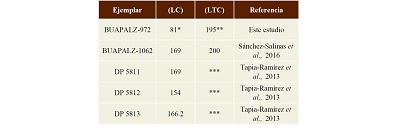
Discusión. El calcáneo de Cuvieronius presenta la tuberosidad alargada y sin protuberancias como en Stegomastodon; la tuberosidad en otros proboscídeos es cuadrangular como en Mammuthus (Oslen, 1960; Tapia-Ramírez et al., 2013; Sánchez-Salinas et al, 2016), el ejemplar BUAPALZ-972 fue comparado con ejemplares adultos de Cuvieronius hyodon de Colima reportados por Tapia-Ramírez (2013) y un ejemplar de Tlaxcala reportado previamente en el área de estudio (Sánchez-Salinas et al., 2016), y se encontró que el ejemplar BUAPALZ-972 presenta similitud en tamaño y forma (Tabla 2); además presenta la tuberosidad alargada y redonda como los ejemplares de Cuvieronius hyodon.
|
Tabla 2. Medidas del ejemplar BUAPALZ-972 y otros ejemplares de calcáneo de Cuvieronius hyodon. Medidas en milímetros (mm).
*ejemplar fragmentado **sin el sustentáculo ***Tapia-Ramírez y colaboradores, no menciona el largo de la tuberosidad del calcáneo (LTC). |
Familia GOMPHOTHERIIDAE Cabrera, 1929
Figura 4b
Material referido. BUAPALZ-969 vértebra.
Descripción. Vértebra lumbar de 22 cm de largo y cuerpo vertebral de 12.5 cm, con margen superior del centro vertebral bilobulado bien definido, mientras que el margen inferior es redondo; lateralmente se puede apreciar el pedículo costal superficial ovalado, corto y cóncavo. El foramen neural es triangular y alargado de 8.2 cm de diámetro; los arcos neurales son bajos y presenta las apófisis articulares. El proceso transversal es corto, robusto y elevado (Figura 4b).
Discusión. Al comparar el ejemplar BUAPALZ-969 con ejemplares de otros proboscídeos se pueden apreciar diferencias en el margen superior del centro vertebral entre las familias de proboscídeos: en los gonfoterios como Stegomastodon y Cuvieronius: el margen ventral superior del centro es bilobulado, bien definido y el margen inferior es redondo; en Mammut, el margen es muy recto, mientras que en Mammuthus se presenta una bilobulación pero no tan definida. Algunos autores, como Boule y Thevenin (1920) o Lucas y colaboradores (2011), mencionan que las vértebras lumbares en gonfoterios como Cuvieronius y Stegomastodon tienen un centro corto, casi circular que carece de facetas para las costillas, también tienten quillas ventrales bajas y superficies laterales cóncavas. Los arcos neurales son bajos y cortos con espinas neurales cortas, inclinadas posteriormente que terminan con espinas expandidas y puntas distales rugosas (Boule y Thevenin, 1920; Lucas et al., 2011); estas características están presentes en el ejemplar BUAPALZ-969. Para Mammut y Mammuthus, Oslen (1972) y Hodgson y colaboradores (2008) mencionan que las vértebras lumbares de ambos animales tienen centros vertebrales pesados, fuertemente construidos y arcos neurales con espinas neurales cortas y gruesas; el borde ventral del centro vertebral, desde una vista anterior, en Mammut es aplastado y cuadrangular, mientras que en Mammuthus el margen es más redondo (Oslen, 1972; Hodgson et al., 2008); también se observa en los ejemplares de Mammuthus que los procesos neurales son elevados, a diferencia de Mammut, en el que los procesos neurales son rectos; los procesos neurales del ejemplar de Tlaxcala son similares a los de Mammuthus.
|
Figura 4. Proboscídeos fósiles adultos y juveniles de San Mateo Huexoyucán, Tlaxcala, México. A) BUAPALZ-972, calcáneo fragmentado de Cuvieronius hyodon; B) BUAPALZ-969, vértebra lumbar de un gonfoterio adulto; C) BUAPALZ-1101; D) BUAPALZ-1102, escápulas juveniles de Mammuthus; E) fíbula fragmentada de Mammuthus. La escala representa 10 cm. Abreviaturas: ACR, acromion; AN, arco neural; BL, bilobulación; CE, cuello de la escapula; CGL, cavidad glenoidea; EPD, epífisis distal; ESN, espina neural; FA, faceta articular; FN, foramen neural; HE, hoja escapular; TBC, tuberosidad calcánea. |

Sin embargo, las características que presentan el ejemplar BUAPALZ-969 como los arcos neurales bajos y cortos, un pedículo costal corto y cóncavo, una bilobulación de la margen ventral bien definida, se le asigna como un gonfoterio, y el tamaño del centro sugiere que corresponda a un ejemplar adulto.
Orden PROBOSCIDEA Illiger, 1811
Familia ELEPHANTIDAE Cabrera, 1929
Mammuthus Brookes, 1828
Mammuthus sp.
Figura 4 c, 4d y 4e
|
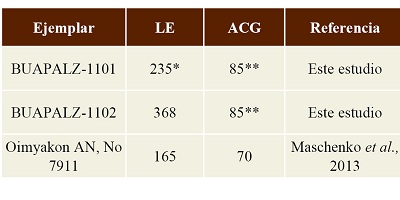
Tabla 3. Medidas de las escápulas BUAPALZ 1101 y BUAPALZ 1102 y la escapula juvenil de Mammuthus reportado por Maschenko et al. (2013). Medidas en milímetros (mm). Longitud escápula= LE, ACG= Ancho Cavidad Glenoidea.
*hoja escapular fragmentada **cavidad glenoidea fragmentada |
Material referido. BUAPALZ-1101, BUAPALZ-1102 escápulas.
Descripción. BUAPALZ-1102 escápula izquierda de 23.5 cm de largo, la hoja escapular rota y tiene un borde redondo; el cuello escapular es estrecho de 11.5 cm, el acromion está fragmentado y se eleva lateralmente y presenta una espina escapular pequeña y fragmentada, la cavidad glenoidea es de 8.5 cm, fragmentada amplia, rectangular y cóncava (Figura 4c).
BUAPALZ-1101 escápula derecha fragmentada de 36.8 cm de largo, hoja escapular completa y con el borde redondo, con el acromion roto y proyectado lateralmente; la cavidad glenoidea de 8.5 cm es amplia, rectangular, cóncava y fragmentada, no conserva la espina escapular, el cuello escapular es estrecho y corto, de 11.5 cm, y presenta un pequeño tubérculo fragmentado (Figura 4d).
Discusión. El tamaño de los ejemplares sugiere que se trata de animales grandes, probablemente proboscídeos, sin embargo, muy pequeños para pertenecer a ejemplares adultos; en Mammuthus el borde es redondo y el cuello de la escapula es corto y estrecho, y la fosa glenoidea es amplia y rectangular (Oslen, 1972), características que están también presentes en los ejemplares de Tlaxcala.
Los ejemplares fueron comparados con ejemplares juveniles de Mammuthus trogontherii reportados por Fisher y colaboradores (Fisher et al., 2014) y Maschenko y colaboradores (2013), en los cuales el acromion se proyecta lateralmente y el cuello de la escápula es estrecho (Fisher et al., 2014) al igual que en los ejemplares estudiados. Se tienen registros de Mammuthus en la localidad de San Mateo Huexoyucán, Tlaxcala, los cuales son ejemplares adultos (Sánchez-Salinas et al., 2016), sin embargo, sólo se cuenta con registros de ejemplares juveniles en Tocuila, Estado de México, para el país (Morett et al., 1998; Arroyo-Cabrales et al., 2002).
Lamentablemente los ejemplares de Tlaxcala no fueron asociados con material más diagnóstico, por ejemplo molares o defensas, para poder calcular la edad biológica del ejemplar, sin embargo los ejemplares de San Mateo Huexoyucán son más grandes en medidas que los ejemplares de crías de Mammuthus reportados por Maschenko y colaboradores (Maschenko et al., 2013), quienes proponen una edad biológica de 14-16 meses de vida al ejemplar de Oimyakon (Tabla 3), lo que sugiere que los ejemplares estudiados son jóvenes.
Material referido. BUAPALZ-965 fíbula.
Descripción. Fíbula fragmentada de 54.5 cm de largo, ancha y estrecha; el ancho de la epífisis distal es de 8.5 cm, la cual es plana fragmentada, terminando en una punta robusta; el maléolo está roto y carece de la epífisis proximal (Figura 4e).
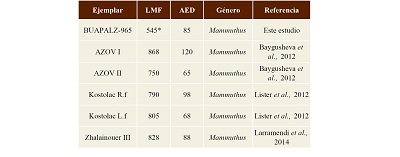
Discusión. El ejemplar de Tlaxcala se distingue de Cuvieronius ya que en este género la epífisis proximal es ancha, plana y redonda (Boule y Thevenin, 1920; Tapia-Ramírez et al., 2013). La fíbula en Mammuthus, en cambio, es más alargada, estrecha y termina en una punta en el extremo de la epífisis distal (Oslen, 1972), característica presente en el ejemplar BUAPALZ-965; al ser comparado con ejemplares de Mammuthus reportados por Baygusheva (2012), Lister (2012) y Larramendi (2014), se encontró que el ejemplar BUAPALZ-965 muestra similitud en el ancho del ancho de la epífisis distal de la fíbula, identificándolo como Mammuthus (Tabla 4).
|
Figura 5. Elementos dentales y postcraneales de équidos pleistocénicos de la Barranca de San Mateo Huexoyucán, Tlaxcala, México. A) BUAPALZ-975, molar inferior de Equus mexicanus; B) BUAPALZ-982; C) BUAPALZ-976, molariformes inferiores de Equus conversidens; D) BUAPALZ-977 vértebra lumbar de Equus sp. En A-C la escala representa 3 cm. En D la escala representa 5 cm. Abreviaturas: APA, apófisis articular; APT, apófisis transversa; CEN, cuerpo vertebral; ECN, entocónido; ESN, espina neural; FN, foramen neural; LFX, linguaflexído; MET, metacónido; MST, metastílido; PLI, plicaballín. |

Orden PERISSODACTYLA Owen, 1848
Familia EQUIDAE Gray, 1821
Equus Linnaeus, 1758
Equus mexicanus Hibbard, 1955
Figura 5
|
Tabla 4. Medidas del ejemplar BUAPALZ-965 y su comparación con la fíbula otros proboscídeos. Medidas en milímetros (mm).
*El ejemplar BUAPALZ-965 está fragmentado, solo conserva la epífisis proximal. |
Material referido. BUAPALZ-975 m1.
Descripción. BUAPALZ-975, molar inferior aislado, robusto, cuadrado, con el borde labial recto y fragmentado de 3 cm de largo; conserva el cemento dental debido a la fragmentación de la corona; no presenta el entocónido, metastílido, metacónido ni el metastílido, sin embargo, conserva el linguafléxido, cuya configuración es en forma de “V”, el pli-caballínido está presente y fuertemente desarrollado (Figura 5a).
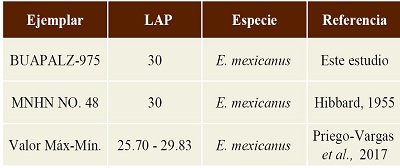
Discusión. La configuración de los linguafléxido de los molariformes inferiores de Equus mexicanus presenta forma de “V” (Hibbard, 1995; Bravo-Cuevas et al., 2011; Alberdi et al., 2014; Priego-Vargas et al., 2017), características presentes asimismo en el molar estudiado. Al comparar el ejemplar BUAPALZ-975 con el de Equus mexicanus reportado por Hibbard (1955), se encontró que el ejemplar de Tlaxcala es similar en medidas al ejemplar proveniente de la Cuenca de México, aunque ambos sobrepasan los valores máximo y mínimo para esta especie reportados por Priego-Vargas y colaboradores (2017), sin embargo Equus mexicanus se caracteriza por ser un équido pleistocénico de talla grande, a decir de Winans, 1989; Bravo-Cuevas et al., 2011; Alberdi et al., 2014; Priego-Vargas et al., 2017, que identifican al ejemplar BUAPALZ-975 como Equus mexicanus (Tabla 5).
Equus conversidens Owen, 1869
Material referido. BUAPALZ-982 m1; BUAPALZ-976 m3.
Descripción. El ejemplar BUAPALZ-982 es un molar inferior aislado de 2.4 cm de largo, fragmentado y sin cemento dental; presenta el entocónido redondo y la configuración del metastílido es aguzada y bien definida, mientras que el metacónido es redondo y la configuración del linguafléxido es en forma de “V”; el pli-caballínido está presente y moderadamente desarrollado (Figura 5b).
BUAPALZ-976 es un molar inferior aislado, de forma triangular y fragmentado, de 2.5 cm de largo, sin cemento dental; el paralófido está fragmentado, el metacónido y entocónido son redondos, el metastílido es aguzado y el linguafléxido tiene forma de “V”; el pliegue del pli-caballínido está presente y moderadamente desarrollado (Figura 5c).
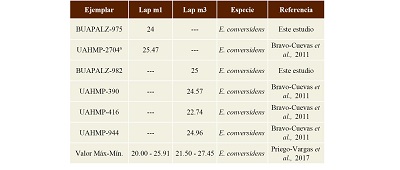
Discusión. En los molares inferiores de Equus conversidens la configuración del linguafléxido predominante es en forma de “V” (Hibbard, 1955; Bravo-Cuevas et al., 2011; Alberdi et al., 2014; Priego-Vargas et al., 2017); se ha considerado a Equus conversidens como un caballo de talla mediana (Bravo-Cuevas et al., 2011; Alberdi et al., 2014; Priego-Vargas et al., 2017). Los ejemplares BUAPALZ-982 y BUAPALZ-976; fueron comparados con ejemplares de Equus conversidens del Estado de Hidalgo reportados por Bravo-Cuevas y colaboradores (2011) y a pesar de la fragmentación y carencia del cemento dental, estos son similares en medidas y también se encuentran dentro de los valores máximos y mínimos para Equus conversidens reportados por Priego-Vargas et al. (2017, Tabla 6).
|
Figura 6. BUAPALZ-1090 Osteodermo aislado de Glyptotherium. La escala representa 4 cm. Abreviaturas: FC, figura central; FP, figura periférica; FPL, folículo piloso. |

La carencia del cemento dental en la mayoría de los molares de équidos pleistocénicos puede ser asociada a la acidez del suelo, lo que propone que la Barranca de San Mateo Huexoyucán ofrece potencial no solo para estudios paleontológicos, sino también para estudios pedoestratigráficos.
Equus sp.
Material referido. BUAPALZ-977, vértebra lumbar.
Descripción. Vértebra aislada de 19.45 cm de largo, con la espina neural fragmentada y el foramen neural elíptico; las apófisis articulares craneales están conservadas, son pequeñas y cóncavas, las caudales son convexas. El cuerpo vertebral completo, de 6.2 cm de largo, es elíptico y la cresta ventral está fragmentada. Las apófisis transversas son planas, fragmentadas y se inclinan dorsoventralmente (Figura 5d).
Discusión. En Equus el cuerpo vertebral de las lumbares es elíptico y presenta crestas ventrales, las apófisis articulares craneales tienen superficies cóncavas y las caudales son convexas, las apófisis transversas son largas y aplanadas dorsoventralmente e inclinadas ligeramente en dirección ventral o dorsal (Sisson y Grossman, 1978); estas características identifican al ejemplar de Tlaxcala como perteneciente a un équido Pleistocénico, ya que en Bison y Bos las vértebras lumbares tienen apófisis espinosas relativamente bajas y anchas (Oslen, 1960; Sisson y Grossman, 1978), característica ausente en el ejemplar BUAPALZ-976.
|
Tabla 5. Medidas del largo anteroposterior del molar inferior del ejemplar BUAPALZ-975 y su comparación con ejemplares de Equus mexicanus. Medidas en milímetros (mm).
|
Orden CINGULATA Illiger, 1811
Familia GLYPTODONTIDAE Gray, 1869
Glyptotherium Osborn, 1903
Glyptotherium cf. floridanum
Material referido. BUAPALZ-1090 osteodermo.
Descripción. Osteodermo aislado de contorno hexagonal de 5.5 cm de largo; su figura central es plana, ligeramente cóncava y abarca la mitad del material referido; el surco marginal es visible con tres folículos pilosos y ocho figuritas periféricas visibles que son más pequeñas que la figura central (Figura 6).
Discusión. Se han registrado tres especies de gliptodontes para el Rancholabreano de México: G. cylindrincum, G. floridanum y G. mexicanum; dos especies reportadas en la Barranca de San Mateo Huexoyucan: Glyptotherium cylindrincum (Ramírez-Cruz y Montellano-Ballesteros, 2014) y Glyptotherium floridanum (Sánchez-Salinas et al., 2016). Ambas especies coexistieron durante el Pleistoceno Tardío (Gillette y Ray, 1981) y han sido reportadas en el centro de México (Arroyo-Cabrales et al., 2008; Bravo-Cuevas et al., 2009; Ramírez-Cruz y Montellanos-Ballesteros, 2014). El ejemplar BUAPALZ-1090 presenta una figura central ligeramente cóncava, al igual que los ejemplares de Glyptotherium floridanum del estado de Hidalgo (Bravo-Cuevas et al., 2009) y de Tlaxcala (Sánchez-Salinas et al., 2016). Las placas de esta especie de gliptodonte tienen figuras centrales aproximadamente iguales que las periféricas, y por lo general son ligeramente levantadas y débilmente cóncavas (Gillette y Ray, 1981; Bravo-Cuevas et al., 2009), mientras que G. cylindricum presenta figuras centrales con una superficie que varía de plana a ligeramente convexa y son más grandes que las periféricas; G. mexicanum se distingue por tener figuras centrales más grandes que las periféricas (Gillette y Ray, 1981; Mead et al., 2007; Ramírez-Cruz y Montellano-Ballesteros, 2014), características ausentes en el material estudiado, al no contar con material más diagnóstico como los molares o el cráneo; se designan al ejemplar BUAPALZ-1090 tentativamente como G. floridanum, siendo el tercer registro de gliptodontes y el segundo registro de esta especie de gliptodonte para San Mateo Huexoyucán, Tlaxcala.
- Discusión
Los ejemplares revisados corresponden a una asociación de mamíferos herbívoros pleistocénicos incluyendo Artiodáctilos (Bison antiquus, Camelops hesternus), proboscídeos (Cuvieronius hyodon, Mammuthus sp.), équidos (Equus conversidens, Equus mexicanus) y gliptodontes (Glyptotherium), los cuales corresponden con una asociación de mamíferos herbívoros rancholabreanos y concuerdan con los registros previos (Bonilla-Toscano, 2011; Ramírez-Cruz y Montellanos-Ballesteros, 2014; Sánchez-Salinas et al., 2016). La combinación de los siete taxa encontrados en este estudio se presenta para los sitios de Chapala-Zacoalco en Jalisco, Tequixquiac en el Estado de México y Valsequillo, Puebla (Ceballos et al., 2010; Ferrusquía-Villafranca et al., 2010), siendo San Mateo Huexoyucán, la cuarta localidad en el país con este ensamble de megafauna (Figura 7). Es interesante observar que esta combinación de megafauna sólo se presenta en la provincia morfotectónica de la Faja Volcánica Transmexicana (Ferrusquía-Villafranca et al., 2010).
|
Figura 7. Localidades del Pleistoceno de México que presentan el mismo ensamble de megafauna (círculos blancos) que la encontrada en San Mateo Huexoyucan (estrella blanca) y localidad con el registro de Mammuthus juvenil (cuadro blanco); todas presentes en la Faja Volcánica Transmexicana (FVTM, área punteada). C-Z, Chapala-Zoacalco, Jalisco; TEQ, Tequixquiac, Estado de México; TOC, Tocuila, Estado de México; SMH, San Mateo Huexoyucan, Tlaxcala; VAL, Valsequillo, Puebla. |
Recientemente se han reportado registros de Mammuthus en México para los estados de Baja California Sur (Rochín-Bañaga et al., 2017), Puebla (Carbot-Chanona et al., 2017) y Oaxaca (Viñas-Vallverdú et al., 2017; Díaz-Sibaja et al., 2018), pero la presencia de organismos juveniles de Mammuthus sólo han sido reportados en Tocuila, Estado de México (Morett et al., 1998; Arroyo-Cabrales et al., 2002), por lo que el registro en San Mateo Huexoyucán sería el segundo reporte para el país de organismos juveniles de Mammuthus (Figura 7). La presencia de ejemplares adultos y juveniles de Mammuthus sugiere el indicio de posibles manadas de mamuts en San Mateo Huexoyucán durante el Pleistoceno de Tlaxcala y estos ejemplares reflejan parte del potencial para estudios paleontológicos y geológicos futuros.
- Conclusión
Se identificaron cuatro órdenes (Artiodactyla, Proboscidea, Perissodcatyla y Xenathra); seis familias (Bovidae, Camelidae, Elephantidae, Equidae, Glyptodontidae y Gomphotheriidae) y ocho especies (Bison antiquus, Camelops hesternus, Cuvieronius hyodon, Equus conversidens, Equus mexicanus, Equus sp., Glyptotherium cf. floridanum y Mammuthus sp.), las cuales concuerdan con la Edad Mamífero-Norteamericana Rancholabreana.
|
Tabla 6. Medidas del Largo anteroposterior de los ejemplares BUAPALZ-975 y BUAPALZ-982 y su comparación con ejemplares de Equus conversidens. Medidas en milímetros (mm).
|

El sitio fósil de San Mateo Huexoyucán es importante por presentar un ensamble de Megafauna sólo encontrado en otras localidades del país dentro de la Faja Volcánica Transmexicana y es el segundo sitio con presencia de juveniles de Mammuthus en México.
La Colección BUAPALZ fue actualizada con nuevos fósiles pleistocénicos como los ejemplares juveniles de mamuts, calcáneos y vertebra de gonfoterios, osteodermos de gliptodontes, molares inferiores y elementos post-craneales de équidos y camélidos, contribuyendo así al registro fósil del Pleistoceno de la Cuenca Puebla-Tlaxcala.
Agradecimientos
Agradecemos a los vecinos y autoridades de San Mateo Huexoyucán, Tlaxcala, nuevamente por su confianza y autorización para el trabajo en campo; al biólogo Luis Bonilla Toscano por la limpieza y revisión de parte del material estudiado; a la Vicerrectoría de Investigación y estudios de Posgrado de la BUAP por el financiamiento de la investigación otorgado al M. en C. Carlos Castañeda-Posadas.
Referencias
Alberdi, M.T., Arroyo-Cabrales, J., Marín-Leyva, A.H., Polaco, O.J., 2014, Study of cedral horses and their place in the Mexican Quaternary: Revista Mexicana de Ciencias Geológicas, 31, 221–237.
Arroyo-Cabrales, J., Morett, L.A., Polaco, O.J., 2002, Tocuila and its research/public outreach program, en Lauwerier y Plug, I. (eds.), The future from the past, 9th ICAZ Conference of International Council of Archaezoology, Durham, editado por R.C.G.M. :153–158.
Arroyo-Cabrales, J., Carreño, A.L., Lozano-García, S., Montellano-Ballesteros, M., 2008, La Diversidad en el pasado, en Soberón, J., Halffter, G., Llorente-Bousquets, J. (comps.), Capital natural de México, Conocimiento actual de la biodiversidad: México, CONABIO: 227–262.
Baygusheva, V.S., Titov, V.V., Timonina, G.L., 2012, Two skeletons of Mammuthus trogontherii from the Sea of Azov Region: Quaternary International, 276, 242–252. https://doi.org/10.1016/j.quaint.2011.05.045
Bell, C.J., Lundelius Jr., E.L., Barnosky, A.D., Graham, R.W., Lindsay, E.H., Ruez Jr., D.R., Semken Jr., H.A., Webb, S.D., Zakrzewski, R.J., 2004, The Blancan, Irvingtonian and Rancholabrean Mammal Ages, en Woodburne, M.O. (ed.)., Late Cretaceous and Cenozoic mammals of North America: United States, Columbia University Press, 232–314. https://doi.org/10.7312/wood13040-009
Bonilla-Toscano, L., 2011, Descripción e identificación de la mastofauna fósil del municipio de Panotla, Tlaxcala: Puebla, BUAP, Escuela de Biología, Tesis de Licenciatura, 52 p.
Boule, M., Thevenin, A., 1920, Mammifères fossiles de Tarija: Paris, Soudier, 256 p.
Bravo-Cuevas, V.M., Ortiz-Caballero, E., Cabral-Perdomo, M.A., 2009, Gliptodontes (Xenarthra, Glyptodontidea) del Pleistoceno Tardío (Rancholabreano) de Hidalgo, centro de México: Boletín de la Sociedad Geológica Mexicana, 61(2), 26–277. https://doi.org/10.18268/bsgm2009v61n2a14
Bravo-Cuevas, V.M., Jiménez-Hidalgo, E., Priego-Vargas, J., 2011, Taxonomía y hábito alimentario de Equus conversidens (Perissodactyla, Equidae) del Pleistoceno Tardío (Rancholabreano) de Hidalgo, centro de México: Revista Mexicana de Ciencias Geológicas, 28, 65–82.
Brookes, J., 1828, A catalogue of the anatomical and zoological museum of Joshua Brookes: London, Taylor, 1, 1–70.
Cabrera, A., 1929, Una revisión de los mastodontes argentinos: Revista del Museo de la Plata, 32, 61-144.
Carbot-Chanona, G., Lagunas-Rodríguez, Z., Jiménez-Moreno, F.J., Suárez, S., 2017, Aspectos paleobiológicos de dos ejemplares de Mammuthus columbi (Mammalia, Proboscidea, Elephantidae) del Pleistoceno de Puebla, centro de México: Boletín de la Sociedad Geológica Mexicana, 69(3), 591–609. https://doi.org/10.18268/bsgm2017v69n3a5
Ceballos, G., Arroyo-Cabrales, J., Ponce, E., 2010, Effects of Pleistocene environmental changes on the distribution and community structure of the mamalian fauna of Mexico: Quaternary Research, 73(3), 464–473. https://doi.org/10.1016/j.yqres.2010.02.006
Díaz-Sibaja, R., Jiménez-Hidalgo, E., García-Zepeda, M.L., 2018, Una nueva localidad fosilífera de Oaxaca (México) y el registro más austral de Bison latifrons. Implicaciones paleobiogeográficas, paleoecológicas y paleoambientales: Boletín de la Sociedad Geológica Mexicana, 70(1), 201–222. https://doi.org/10.18268/bsgm2018v70n1a12
Donovan, S.K. y Paul, C.R.C., 1998, The adequacy of the fossil record: Chichester, John Wiley and Sons, 312 p.
Fariña, R.A., Vizcaino, S.F., De Iuliis, G., 2013, The Great American Biotic Interchange. Megafauna: Giant beasts of Pleistocene South America: Bloomington, Indiana University Press, 150 p.
Ferrusquía-Villafranca, I., Arroyo-Cabrales, J., Martínez-Hernández, E., Gama-Castro, J., Ruiz-González, J., Polaco, O.J., Johnson, E., 2010, Pleistocene mammals of Mexico: A critical review of regional chronofaunas, climate change response and biogeographic provinciality: Quaternary International, 217(1-2), 53-104. https://doi.org/10.1016/j.quaint.2009.11.036
Ferrusquía-Villafranca, I., Ruiz-González, J., Martínez-Hernández, E., Gama-Castro, J., de Anda-Hurtado, P., Polaco, O.J., 2017, Quaternaty mammals, people and climate change: A view from Southern North America, en Monks, G.G. (ed.)., Climate change and human responses: A zooarchaeological perspective: Vertebrate Paleobiology and Paleoanthropology, 27-64 https://doi.org/10.1007/978-94-024-1106-5_3
Fischer, G., 1814, Zoognosia. Tabulis synopticis illustrate: Moscow, Typis Nicolai Sergeidis Vsevolozsky, 3, 1–694.
Fisher, D.C., Shirley, E.A., Whalen, C.D., Calamari, T.Z., Rountrey, A.N., Tikhonov, A.N., Buigues, B., Lacombat, F., Grigoriev, S., and Lazarev, P.A., 2014, X-Ray computed tomography of two mammoth calf mummies: Journal of Paleontology, 88 (4) 664-675. https://doi.org/10.1666/13-092
France, D.L., 2009, Human and no human bone identification: a color atlas: Boca Ratón, Florida, Taylor & Francis Group, 1–773 p. https://doi.org/10.1201/9781420062878
Galotta, D.R. y Galotta, J.M., 1988, Osteología de la llama (Lama glama guanicoe f.d. glama L. 1758): Revista de Ciencias Agrarias de la Facultad de Ciencias Agrarias, IX (1/43), 19–61.
Gillette, D.D. y Ray, C.E., 1981, Glyptodonts of North America: Smithsonian Contributions to Paleobiology, 40, 1–255. https://doi.org/10.5479/si.00810266.40.1
Gómez-Pérez, L.E. y Carbot-Chanona, G., 2012, Contribución al estudio de los megamamíferos del Pleistoceno Tardío del municipio de Villaflores, Chiapas, México: Lacandonia, 6, 31-44.
Gray, J.E., 1821, On the natural arrangement of vertebrose animals: London Medical Repository Review, 15, 296–310.
Gray, J.E., 1869, Catalogue of carnivorous, pachydermatous, and dentate mammalia in the British Museum: London, British Museum Natural History, 398 p.
Hibbard, W.C., 1955, Pleistocene vertebrates from the Upper Becerra (Becerra Superior) Formation, Valley of Tequixquiac, Mexico, with notes on other Pleistocene forms: Contributions from the Museum of Paleontology, University of Michigan, 21(5), 47–96.
Hillson, S., 2005, Teeth: Cambridge, Cambridge University Press (Cambridge Manuals in Archaeology), 373 p.
Hodgson, J.A., Allmon, W.D., Nester, P.L., Sherpa, J.M., Chiment, J.J., 2008, Comparative osteology of late Pleistocene mammoth and mastodon remains from the Watkins Glen site, Chemung County, New York, en Allmon, W.D., Peter, L.N. (eds.), Mastodon Paleobiology, taphonomy, and paleoenvironment in the late Pleistocene of New York State: Studies on the Hyde Park, Chemung, and North Java Sites: Palaeontographica Americana, 61, 301–367.
Illiger, C., 1811, Prodromus systematis mammalium et avium additis terminis zoographicis utriusque classis, eorumque versione germanica (en línea): Berlin, Salfeld, 1, 1‒301. http://dx.doi. org/10.5962/bhl.title.106965
Instituto Nacional para el Federalismo y el Desarrollo Municipal, 2010, Enciclopedia de los municipios y delegaciones de México, Estado de Tlaxcala, disponible en <http://www.inafed.gob.mx/work/enciclopedia/EMM29tlaxcala/index.html.>, consultado el 16 de julio de 2018.
Jiménez-Hidalgo, E., Cabrera-Pérez, L., MacFadden, B.J., Guerrero-Arenas, R., 2013, First record of Bison antiquus from the Late Pleistocene of southern Mexico: Jounal of South American Earth Sciences, 42, 83–90. https://doi.org/10.1016/j.jsames.2012.07.011
Larramendi, A., 2014, Skeleton of a Late Pleistocene steppe mammoth (Mammuthus trogontherii) from Zhalainouer, Inner Mongolian Autonomus Region, China: Paläontologische Zeitschrift, 89 (2), 229–250. https://doi.org/10.1007/s12542-014-0222-8
Leidy, J., 1852, Memoir on the extinct species of American ox: Smithsonian Contributions. Knowledge, 5 (3), 1‒20. https://doi.org/10.5962/bhl.title.31716
Leidy, J. 1854, Description of a fossil apparently indicating an extinct species of the camel tribe. Proceedings of the Academy of Natural Sciences of Philadelphia, 7, 172–173.
Leidy, J., 1873, Contributions to the extinct vertebrate fauna of the western Territories: United States geological and geographical survey of the Territories, 1, 14‒358.
Linnaeus, K., 1758, Systema naturae perregnatria naturae, secundum classes, ordines, genera, species cum characteribus, differentiis, synonymis, locis: Stockholm, Laurentii Salvii, 824 p.
Lister, A.M., Dimitrijevic, V., Markovic, Z., Knezevic, S., Mol, D., 2012, A skeleton of ‘steppe’ mammoth [Mammuthus trogontherii (Pohlig)] from Drmno, near Kostolac, Serbia: Quaternary International, 276–277, 129–144. https://doi.org/10.1016/j.quaint.2012.03.021
Lucas, G.S., Aguilar, R.H., Spielmann, J.A, 2011, Stegomastodon (Mammalia, Proboscidea) from the Pliocene of Jalisco, Mexico and the species-level taxonomy of Stegomastodon: Bulletin of the New Mexico Museum of Natural History and Science, 53, 517–553.
Lundelius, E., 1972, Fossil vertebrates from the Late Pleistocene Ingleside Fauna, San Patricio County: Texas, Bureau of Economic Geology, University of Texas of Austin, Report of investigations No. 77, 82 p.
Maschenko, E.N., Boeskorov, G.G., Baranov, V.A., 2013, Morphology of a mammoth calf (Mammuthus primigenius) from Ol`chan (Oimiakon, Yakutia): Paleontological Journal, 47 (4), 425–438. https://doi.org/10.1134/s0031030113040096
Mead, J.I., Swift, S.L., White, R.S., McDonald, H.G, Báez, A., 2007, Late Pleistocene (Rancholabrean) Glyptodont and Pampathere (Xenarthra, Cingulata) from Sonora, Mexico: Revista Mexicana de Ciencias Geológicas, 24, 439‒449.
Morett, L., Arroyo-Cabrales, J., Polaco, O.J., 1998, El sitio paleontológico de Tocuila: Arqueología Mexicana, 30, 57.
Olsen, S. J. 1972, The American mastodon and the woolly mammoth. Osteology for the archaeologist. Papers of the Peabody Museum of Arcaheology and Ethnology, Harvard University, 56(3), 1-43.
Osborn, H.F., 1903, Glyptotherium texanum, a new glyptodont, from the Lower Pleistocene of Texas: Bulletin of the American Museum of Natural History, 19, 491–494.
Osborn, H.F., 1923, New subfamily, generic, and specific stages in the evolution of the Proboscidea: American Museum Novitates, 99, 1–4.
Oslen, J.S., 1960, Post-cranial skeletal characters of Bison and Bos: Cambridge, Papers of the Peabody Museum of Archaeology and Ethnology Harvard University, 15 p.
Owen, R., 1848, Description of the teeth and portions of jaws of two extinct anthracotheroid quadrupeds discovered by the Marchioness of Hastings in the Eocene deposits of the N.W. coast of the Isle of Wight: with an attempt to develop Cuvier’s idea of the classification of Pachyderms by the number of their toes: Quarterly Journal Geological Society of London, 4 (1-2), 103‒141. https://doi.org/10.1144/gsl.jgs.1848.004.01-02.21
Owen, R., 1869, On Fossil Remains of Equines from Central and South-America referable to Equus conversidens Ow., and E. arcidens Ow: Philosophical Transactions of the Royal Society of London, 159, 559‒73. https://doi.org/10.1098/rstl.1869.0020
Priego-Vargas, J., Bravo-Cuevas, V.M., Jiménez-Hidalgo, E., 2017, Revisión taxonómica de los équidos del Pleistoceno de México con base en la morfología dental: Revista Brasileira de Paleontologia, 20 (2), 239-268. https://doi.org/10.4072/rbp.2017.2.07
Ramírez-Cruz, G.A., Montellano-Ballesteros, M., 2014, Two new Glyptodont records (Mammalia: Cingulata) from the Late Pleistocene of Tamaulipas and Tlaxcala, Mexico: Implications for the taxonomy of the genus Glyptotherium: The Southwestern Naturalist, 59(4), 522‒530. https://doi.org/10.1894/jkf-45.1
Rochín-Bañaga, H., Schwennicke, T., Herrera-Gil, L.A., 2017, Registro de Mammuthus columbi en el área de El Carrizal, Baja California Sur, México: Boletín de la Sociedad Geológica Mexicana, 69(2), 495–503. https://doi.org/10.18268/bsgm2017v69n2a12
Sánchez-Salinas, M., Jiménez-Hidalgo, E., Castañeda-Posadas, C., 2016, Mamíferos fósiles del Pleistoceno tardío (Rancholabreano) de San Mateo Huexoyucán, Tlaxcala, México: Boletín de la Sociedad Geológica Mexicana, 68(3), 497–514. https://doi.org/10.18268/bsgm2016v68n3a7
Schmid, E., 1972, Atlas of animal bones: for prehistorians and Quaternary geologist: Amsterdam, Elsevier, 153 p.
Sisson, J.D., Grossman, R.G., 1978, Anatomía de los animales domésticos: Madrid, Salvat Editores, t. 1, 1335 p.
Smith, C.H., 1827, Supplement to the order Ruminantia, en Cuvier, G., The animal kingdom arranged in conformity with its organization: London, Geo. B. Whittaker, 4, 33–428.
Stuart, A.J., 1991, Mammalian extinctions in the Late Pleistocene of northern Eurasia and north America: Biological Reviews, 66 (4), 435-562. https://doi.org/10.1111/j.1469-185x.1991.tb01149.x
Tapia-Ramírez, G., Guzmán, A.F., Polaco, J.O., 2013, Los Gonfoterios (Proboscidea, Gomphoteriidae) de Colima, México: Boletín de la Sociedad Geológica Mexicana, 65(3), 591‒607. https://doi.org/10.18268/bsgm2013v65n3a12
Viñas-Vallverdú, R., Arroyo-Cabrales, J., Rivera-González, I.I., Rodríguez-Álvarez, X.P., Rubio-Mora, A., Eudave-Eusebio, I.N., Solís-Torres, O.R., Ardelean, C.F., 2017, Recent archaeo-paleontological finding from Barranca del Muerto site, Santiago Chazumba, Oaxaca, Mexico: Quaternary International, 431, 168–180. https://doi.org/10.1016/j.quaint.2015.04.055
Webb, S.D., 1965, The osteology of Camelops: Los Angeles County Museum Bulletin, 1, 1–54.
Winans, M.C., 1989, A quantitative study of North American fossil species of the genus Equus, en D.R., Prothero & R.M. Soch (eds.), The evolution of Perissodactyls: Oxford University Press, p. 262–297.
Manuscrito recibido: Abril 16, 2018.
Manuscrito corregido: Agosto 07, 2018.
Manuscrito aceptado: Agosto 10, 2018.