
|
Boletín de la Sociedad Geológica Mexicana Volumen 71, núm. 3, 2019, p. 841-849 http://dx.doi.org/10.18268/BSGM2019v71n3a13
|
 |
Short note
Schlueterella stinnesbecki n. sp. (Ammonoidea, Diplomoceratidae) from the Turonian–Coniacian of northeastern Mexico
Christina Ifrim
Institut für Geowissenschaften, Universität Heidelberg, Im Neuenheimer Feld 234, 69120 Heidelberg, Germany.
This email address is being protected from spambots. You need JavaScript enabled to view it.
Abstract
A new species of the genus Schlueterella Wiedmann, 1962 is here described. Twenty specimens of this heteromorph ammonoid were collected in northern Coahuila, Mexico. Schlueterella stinnesbecki n. sp. seems to have been an endemic form in the late Turonian–early Coniacian. This is the oldest definite record of the genus and a basal form in its evolution away from Neocrioceras.
Keywords: Ammonoid, Cretaceous, Turonian, Coniacian, Mexico, endemic.
Resumen
Se describe una nueva especie del género Schlueterella Wiedmann, 1962. Los veinte ejemplares de este ammonoideo heteromorfo fueron colectados en el norte de Coahuila, México. Parece que Schlueterella stinnesbecki n. sp. fue una especie endémica en el Turoniano tardío–Coniaciano temprano. Con total certeza se trata del registro más antiguo de este género y una forma básica en su evolución desde Neocrioceras.
Palabras clave: Ammonoideo, Cretácico, Turoniano, Coniaciano, México, endémico.
1. Introduction
Schlueterella Wiedmann, 1962 is a little known genus, the affinities of which are still debatable. The specimens described here were discovered in northeastern Mexico in the years 2002–2006 and 2018. They originated from two quarries in the quarry area of the municipality of Múzquiz, northwest Coahuila. The Rosario section (R, 28°52.586ʹN, 102°24.222ʹW, Figure 1) comprises the Turonian–Coniacian boundary from the Mytiloides scupini Zone to the upper Cremnoceramus crassus inconstans Zone (Stinnesbeck et al., 2005; Ifrim et al., 2014), whereas the Carranza section (CA, 29°10.775ʹN, 102°27.193ʹW, Figure 1) is much shorter and comprises only the middle part of the lower Coniacian Cremnoceramus crassus inconstans Zone (Ifrim et al., 2011). In both quarries, Schlueterella stinnesbecki n. sp. is a common species.
|
Figure 1. Map of Mexico with the state of Coahuila in gray (below), and zoom on northern Coahuila with location of the Rosario and Carranza quarries (above). |
2. Systematic Paleontology
Higher systematics follows Wright et al. (1996) except for the subfamily and genus for reasons discussed below. Terminology for tubercles, nodes, and spines follows Ifrim et al. (2018).
Repository is the Paleontological Collection (Colección de Paleontología, CPC), housed in the Desert Museum (Museo del Desierto), Saltillo, Coahuila. For lithostratigraphy see Stinnesbeck et al. (2005). For the Rosario section see Ifrim et al. (2014). For the Carranza section and its correlation see Nyborg et al. (2014) and Ifrim et al. (2011). Further abbreviations are D: diameter, WH: whorl height, WI: Distance between whorl at D and previous whorl, U: umbilicus.
Suborder Ancyloceratina Wiedmann, 1966
Superfamily Turrilitoidea Gill, 1871
Family Diplomoceratidae Spath, 1926
Subfamily Diplomoceratinae Spath, 1926
Genus Schlueterella Wiedmann, 1962
Type species. Ancyloceras pseudoarmatum Schlüter, 1872 by original designation of Wiedmann (1962, p. 205). The genus is characterized by an open coil, tubercles from simple or double ribs always on the primaries, if not all ribs bear tubercles.
Originally, Schlueterella was introduced as subgenus of Neocrioceras by Wiedmann (1962). The type species of Neocrioceras is Crioceras spinigerum Jimbo, 1894, (pl. 8, figure 1) sensu Diener, 1925, (p. 192). Neocrioceras was erected by Spath (1921) on the basis of a juvenile whorl of Crioceras spinigerum Jimbo, 1894 (= Schlueterella compressa sensu Klinger, 1976). An emended description of this species was given by Diener (1925). In Neocrioceras, juvenile whorls are moderately helicoid, later more or less crioconic with circular to depressed section, dense straight to prorsiradiate ribs, some with lateral tubercles and ventrolateral tubercles or nodes, the latter opposite or alternating on venter. According to Wiedmann (1962), Schlueterella differs from Neocrioceras in having corresponding instead of alternating ribs and in having more complex instead of simple ribbing.
Wright et al. (1996) placed the subgenus Schlueterella under the genus Pseudoxybeloceras, a view rejected here based on the overall morphology of more complete specimens that show a coiling pattern that matches Neocrioceras, and because Schlueterella is interpreted here to have evolved from Neocrioceras. It may be justified to place some fragmented specimens with straight shafts in the genus Pseudoxybeloceras.
The specimens described here match the original definition of Schlueterella of Wiedmann (1962). In addition, they have a crioconic shell, but with small portions of straighter coiling, but lacking straight shafts as in the Polyptochoceratinae. Matsumoto and Miyauchi (1984) raised Schlueterella to genus level, a view followed by Klinger et al. (2007) and Summesberger et al. (2017), among others. This genus is further interpreted here to have evolved from Neocrioceras and thus included into the subfamily Diplomoceratinae.
|
Figure 2. Schlueterella stinnesbecki sp. nov. from Rosario (R) and Carranza (CA) in northern Coahuila. 1: CPC–853, R, Yellow Member; 2: CPC–914, R40-42; 3: CPC–915, R39-41; 4: CPC–916, R40-42; 5: CPC–917, CA4-5; 6: CPC–918, CA, Yellow Member; 7: CPC–919, R40-42; 8: CPC–920, CA, Yellow Member; 9: CPC–921; 10: CPC–922, 11: CPC–923, 12: CPC–924, 9–12: R, Yellow Member. Scale: 50 mm. |

Schlueterella stinnesbecki n. sp.
Figure 2.1–2.12, Figure 3.1–3.5, Figure 4
2005 ? Neocrioceras sp.; Stinnesbeck et al., fig. 8f
2007 ? Neocrioceras sp.; Ifrim et al., fig. 6d
Diagnosis. Crioconic, in the juvenile stage moderately helicoid shell with up to 38 rursiradiate, simple primary ribs per whorl bearing lateral tubercles and ventrolateral nodes or spines without looping, intercalated regularly by one simple, parallel secondary rib in the juvenile stage and to four in the adult stage. The transition from juvenile to adult ornament is either at D around 50 or around 100 mm, indicative of size dimorphism.
|
Figure 3. Schlueterella stinnesbecki ssp. nov. from the Yellow Member of Rosario (R) and Carranza (CA) in northern Coahuila. 1: CPC–925, R; 2: CPC–927, CA; 3: CPC–989, R; 4: CPC–926, CA; 5: holotype, CPC–990, R. Scale: 50 mm. |

Etymology. This species is named in honor of Wolfgang Stinnesbeck for his extensive paleontological work in Latin America since 1990, including a series of pioneer projects in northeastern Mexico.
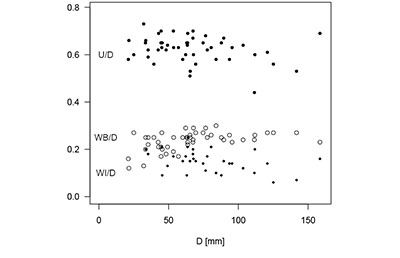
Description. Juvenile shell is crioconic to moderately helicoid and slightly irregular (Figure 2), adult crioconic (e.g., Figure 3.5). Juvenile whorls are delicate with a WH/D of 0.13 to 0.25, at D > 90 mm more stout and regular with a WH/D of 0.25 to 0.30 (Figure 4). Irregular coiling causes variation in WI/D between 0.09 and 0.21 in juvenile whorl and decreases to 0.06 to 0.16 at D > 90 mm. U/D decreases slightly from 0.58–0.73 at small D to 0.53–0.63 at D > 90 mm.
Ornament consists of up to 38 primary ribs per whorl which are straight to prorsiradiate near the dorsum and flex back umbilicolaterally in a wide arch to become rursiradiate towards the venter. They wear tubercles on midflank and ventrolateral nodes or spines. Primary ribs are closer in the juvenile stage, and one to two simple secondaries are intercalated. This number increases to 2–4 simple secondaries at D between 50 and 100 mm, their number increasing with growth. They parallel the primaries. Suture not preserved.
|
Figure 4. Morphometric relationships of Schlueterella stinnesbecki n. sp. during ontogenesis. |
Types. The holotype is CPC–990 (Figure 3.5, also figured in Stinnesbeck et al., 2005, Figure 8f) from an unknown bed in the Yellow Member of the Rosario quarry. Paratypes are CPC–853, CPC–914 to –927 and CPC–989 from the from the uppermost Turonian (Cremnoceramus waltersdorfensis W. Zone) to lower Coniacian (C. crassus inconstans Zone), i.e., the upper Gray Member and the Yellow Member of the Rosario quarry, and the Yellow Member in the Carranza quarry.
Dimensions. See Table 1.
|
Table 1. Morphometry of n. sp.
|

Occurrence. Schlueterella stinnesbecki sp. nov. is only known from northwest Coahuila. It was recorded from the uppermost Turonian (waltersdorfensis Zone) to lower Coniacian (inconstans Zone), beds R24–R42 of the Gray and Yellow members of the Rosario quarry and beds CA1 to CA6 of the Carranza quarry (compare Ifrim et al., 2011, with Ifrim et al., 2014 for correlation).
Discussion. Wiedmann (1962) differentiated two groups within Schlueterella. The group around Sch. pseudoarmatum (Schlüter, 1872) bears four rows of tubercles on primary double ribs, whereas the group around Sch. kossmati (Simionescu, 1899) bears tubercles on simple primaries, or all ribs bear tubercles. The specimens described here clearly belong to the group around Sch. pseudoarmatum (Schlüter, 1872). This type species from the upper Campanian of Spain differs from Sch. stinnesbecki n. sp. by bearing nodes on all ribs and by a straight shaft at a WH of 245 mm.
Sch. tenuiannulatum Collignon, 1969 from the lower Campanian of Madagascar differs in having much finer ribs at WH around 18 mm. Sch. denseornatum Collignon, 1969 from the lower Campanian of Madagascar is has a straight shaft at WH around 13 mm and also has much finer ribs than the material presented here.
Sch. paderbornense (Schlüter, 1872) from the lower Coniacian of Germany differs from Sch. stinnesbecki n. sp. in irregular ribbing and in secondaries which efface on the flank, whereas they cross the dorsum in the forms from Mexico. The specimens described here are closest to Sch. compressus Klinger, 1976 from the middle Coniacian to basal Campanian of South Africa, Madagascar, central and western Europe, Wyoming, California, and Japan (e.g., Klinger and Kennedy, 2003), but this species has maximum 2–3 secondaries between the primary looped ribs, up to three ribs are joined by tubercles, in some specimens tuberculate and non-tuberculate ribs are equally sized. Tubercles are situated on the dorsal third of the flank (Kennedy and Cobban, 1991) instead on mid-flank. In addition, straight sections are absent in the shell of Sch. stinnesbecki n. sp., but it has substraight parts.
Schlueterella sp. from the middle to upper Coniacian of Wyoming (Kennedy and Cobban, 1991) has broad swelling with up to three looped ribs joining at nodes, and up to six intercalated secondaries. In Sch. sp. and N. (Sch.) compressus Klinger, 1976, the presence of ventrolateral nodes indicate formation of spines (compare Ifrim et al., 2018). Both species may have evolved from Sch. stinnesbecki n. sp.
Neocrioceras undulosum Matsumoto, 1977 from the middle? Turonian of Japan has a comparable irregular coiling, ribs and nodes at a D of 40 mm but has fine weak riblets on the ribs and interspaces instead of secondaries. It also bears similarities with Hyphantoceras and Madagascarites, and its generic assignation remains unclear (Matsumoto, 1977; Wiese, 2000).
In Schlueterella cf. maderi Immel, Klinger & Wiedmann, 1982, all ribs loop between tubercles. Sch. spinigerum (Jimbo, 1894) from the Santonian of Japan is more robust and more densely coiled with a higher WH/D and a lower WI/D.
In the specimens described here, the change from the irregularly coiled, more coarsely ornamented juvenile growth stage to the regularly coiled, more delicately ornamented adult growth stage is either at a D around 50 mm or D around 100 mm. This may indicate size dimorphism. It is not noted in the morphometric data (Figure 4) and only visible in the ornamentation.
3. Conclusions
The record of Schlueterella stinnesbecki n. sp. described here is one of the oldest records of the genus . Bed R24 in the Rosario quarry falls into the uppermost Turonian, whereas N. paderbornense is lower Coniacian (Wiese, 2000), and the taxonomic position of Neocrioceras undulosum Matsumoto, 1977 from the middle? Turonian of Japan seems transitional. All other records are younger.
The size dimorphism suggested here has not been recognized before, mostly due to findings and descriptions of isolated and/or incomplete specimens elsewhere.
Ornamentation in Schlueterella stinnesbecki n. sp. is simple compared to later forms which show an increased differentiation in tuberculation and ribbing, i.e. towards finer ribbing, effacing secondaries or increasing number of secondaries. In addition, the substraight parts of some shells show a transition to later species of Schlueterella which have straight parts in their shell. Schlueterella stinnesbecki n. sp. is thus considered a basal form in the evolution of this genus.
Acknowledgements
I thank Wolfgang Stinnesbeck for introducing me to the geology and paleontology of Mexico, for many field campaigns, and for fruitful discussions during the last 20 years. Arturo H. González González is thanked for logistic support, José Manuel Padilla both Desert Museum (Museo del Desierto), Saltillo, Coahuila, Mexico) for access to the CPC collection, Melanie Kling (Heidelberg) for help in field, reviewers Herbert Summesberger (Vienna) and Josep Moreno Bedmar (Mexico City) for their constructive reviews, Griselda Guerrero Márquez and Tom Musselman for editing, and editor Francisco Vega Vera. Financial support of the German Science Foundation (DFG project IF61/11-1) is gratefully acknowledged.
References
Collignon, M., 1969, Atlas des fossiles caractéristiques de Madagascar (Ammonites), Fascicule 15 (Campanien inférieur). Tananarive, Service Géologique, 216 p.
Diener, C., 1925, Ammonoidea neocretacea: Berlin, W. Junk, Fossilum catalogus (1: Animalia), 29, 244 p.
Gill, T., 1871, Arrangement of the Families of the Mollusks: Smithsonian Miscellaneous Collections, 227, xvi+49 p.
Ifrim, C., P. Bengtson, and G. Schweigert, 2018, Growth and function of spines in Jurassic and Cretaceous ammonites: Cretaceous Research, 88, 62–78. https://doi.org/10.1016/j.cretres.2017.05.003
Ifrim, C., W. Stinnesbeck, and E. Frey, 2007, Upper Cretaceous (Cenomanian–Turonian and Turonian–Coniacian) open marine plattenkalk deposits in NE Mexico: Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 245(1), 71–81. https://doi.org/10.1127/0077-7749/2007/0245-0071
Ifrim, C., F. J. Vega, and W. Stinnesbeck, 2011, Epizoic stramentid cirripedes on ammonites from Mexican late Cretaceous platy limestones: Journal of Paleontology, 85(3), 526–538. https://doi.org/10.1666/10-052.1
Ifrim, C., F. Wiese, and W. Stinnesbeck, 2014, Inoceramids and biozonation across the Turonian-Coniacian boundary (Upper Cretaceous) at El Rosario, Coahuila, northeastern Mexico: Newsletters on Stratigraphy, 47(2), 211–246. https://doi.org/10.1127/0078-0421/2014/0043
Immel, H., H. C. Klinger, and J. Wiedmann, 1982, Die Cephalopoden des Unteren Santon der Gosau von Brandenberg/Tirol, Österreich: Zitteliana, 8, 3–32.
Jimbo, K., 1894, Beiträge zur Kenntnis der Fauna der Kreideformation von Hokkaido: Paläontologische Abhandlungen, Neue Folge, 2, 140–198.
Kennedy, W. J., and W. A. Cobban, 1991, Coniacian ammonite faunas from the United States Western Interior: Special Papers in Palaeontology, 45, 96 p.
Klinger, H. C., 1976, Cretaceous heteromorph ammonites from Zululand: Memoirs of the Geological Survey of the Republic of South Africa, 69, 142 p.
Klinger, H. C., and W. J. Kennedy, 2003, Cretaceous faunas from Zululand and Natal, South Africa, The ammonite families Nostoceratidae Hyatt, 1894 and Diplomoceratidae Spath, 1926: Annals of the South African Museum, 110(6), 219–334.
Klinger, H. C., W. J. Kennedy, and W. E. Grulke, 2007, New and little-known Nostoceratidae and Diplomoceratidae (Cephalopoda: Ammonoidea) from Madagascar: African Natural History, 3, 89–115.
Matsumoto, T., 1977, Some heteromorph ammonites from the Cretaceous of Hokkaido: Memoirs of the Faculty of Science, Kyushu University, Series D, Geology, 23(3), 303–366.
Matsumoto, T., T. Miyauchi, 1984, Some Campanian ammonites from the Soya area, in T. Matsumoto (ed.), Some Ammonites from the Campanian (Upper Cretaceous) of Northern Hokkaido. Palaeontological Society of Japan, Special Paper: Special Papers of the Paleontological Society of Japan, 27, 33–76.
Nyborg, T., C. Ifrim, J. A. Moreno-Bedmar, H. Porras Muzquiz, S. Giersch, and F. J. Vega, 2014, Late Cretaeous fish cans: Fish preserved in ammonite body chambers from the middle Santonian of Coahuila State, northeastern Mexico: Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 273(1), 75–88. https://doi.org/10.1127/0077-7749/2014/0417
Schlüter, C., 1872, Cephalopoden der oberen deutschen Kreide I: Palaeontographica, 21, 1–120.
Simionescu, J., 1899, Fauna cretacica superióra de la Ürmös (Transilvania): Academia Romana, Publicatiunile Fondului Vasilie Adamachi, 4(1), 239–275.
Spath, L. F., 1921, On Upper Cretaceous Ammonoidea from Pondoland: Annals of the Durban Museum, 3(2), 39–57.
Spath, L. F., 1926, On new ammonites from the English Chalk: Geological Magazine, 63(2), 77–83. https://doi.org/10.1017/s0016756800083710
Stinnesbeck, W., C. Ifrim, H. Schmidt, A. Rindfleisch, M.-C. Buchy, E. Frey, A. H. González González, F. J. Vega-Vera, H. Porras-Muzquiz, L. Cavin, G. Keller, and K. T. Smith, 2005, A new lithographic limestone deposit in the Upper Cretaceous Austin Group at El Rosario, county of Muzquiz, Coahuila, northeastern Mexico: Revista Mexicana de Ciencias Geológicas, 22(3), 401–418.
Summesberger, H., W. J. Kennedy, and P. Skoumal, 2017, Early and middle Santonian Cephalopods from the Gosau Group (Upper Cretaceous, Austria) 2. Heteromorph Ammonoidea: Abhandlungen der Geologischen Bundesanstalt, 71, 101–149.
Wiedmann, J., 1962, Ammoniten aus der Vascogotischen Kreide (Nordspanien) I. Phylloceratina, Lytoceratina: Palaeontographica, 73, 49–104.
Wiedmann, J., 1966, Stammesgeschichte und System der posttriadischen Ammonoideen: ein Überblick: Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 125(1–3), 49–78.
Wiese, F., 2000, On some late Turonian and early Coniacian (Upper Cretaceous) heteromorph ammonites from Germany: Acta Geologica Polonica, 50(4), 407–419.
Wright, C. W., J. H. Calloman [Sic], and M. K. Howarth, 1996, Cretaceous Ammonoidea, in R. L. Kaesler (ed.), Treatise on Invertebrate Paleontology:Part L, Mollusca 4, revised, Boulder, Lawrence, University of Kansas & Geological Society of America, xx+362 p.
Manuscript received: April 6, 2018.
Corrected manuscript received: July 1, 2018.
Manuscript accepted: July 10, 2018.