
|
BOLETÍN DE LA SOCIEDAD GEOLÓGICA MEXICANA Vol 65, Núm. 3, 2013, P. 581-590 1 http://dx.doi.org/10.18268/BSGM2013v65n3a11 |
 |
Chancelloriida Walcott, 1920 y Reticulosa Reid, 1958 del Cámbrico medio de San José de Gracia, Sonora, México
Francisco J. Cuen1, M.S. Beresi2, A. Montijo1, B.E. Buitrón3,*, I. Minjárez1, M. de la O1, J. J. Palafox1
1 Universidad de Sonora, División de Ciencias Exactas y Naturales, Departamento de Geología, Blvd. Luis Encinas y Rosales, CP 83000, Hermosillo, Sonora.
2 Instituto Argentino de Nivología, Glaciología y Ciencias Ambientales (IANIGLA) Consejo Nacional de Investigaciones Científicas y Tecnológicas (CONICET), Av. A. Ruíz Leal s/n, Parque General San Martín, 5500, Mendoza, Argentina.
3 Universidad Nacional Autónoma de México, Instituto de Geología, Departamento de Paleontología, Ciudad Universitaria, CP 04510, México D.F., México
* This email address is being protected from spambots. You need JavaScript enabled to view it.
Resumen
Se describen escleritos aislados de Chancelloria eros Walcott, 1920, Diagoniella sp. y asociaciones de espículas hexactinélidas-estauractinas y hexactina-pentactinas, para el Cámbrico medio de San José de Gracia, Sonora, México; el material procede de una secuencia sedimentaria marina de plataforma, constituida principalmente por caliza intercalada con lutita, en asociación con fósiles representativos de la Biozona de Albertella, como son Peronopsis bonnerensis Resser, 1938; Oryctocephalites walcotti Reeser, 1938; Elrathina antiqua Palmer y Halley, 1979 y Ogygopsis typicalis Resser, 1939, además de braquiópodos inarticulados.
Chancelloria eros Walcott, 1920 ha sido reportada anteriormente para el Cámbrico medio de la Formación Arrojos, así como Chancelloria sp. para el Cámbrico Inferior de la Formación Puerto Blanco, en asociación con ?Kiwetinokia del Cerro Rajón situado en la región de Caborca, lo cual constituye el primer registro para San José de Gracia, Sonora central.
Palabras clave: Chancelóridos, Protospóngidos, Cámbrico, Sonora, México.
Abstract
Isolated sclerites of Chancelloria eros Walcott, 1920, Diagoniella sp. and associations of stauractine-hexactinellid and hexactine-pentactine spicules are described for the middle Cambrian of San José de Gracia, Sonora, Mexico; the material proceeds from a shelf marine sedimentary sequence, consisting mainly of shale interbedded with limestone, in association with fossils representatives of the Biozone Albertella, like Peronopsis bonnerensis Resser, 1938; Oryctocephalites walcotti Reeser, 1938; Elrathina antiqua Palmer and Halley, 1979, Ogygopsis typicalis Resser, 1939, and inarticulate brachiopods.
Chancelloria eros Walcott, 1920 has been previously reported for middle Cambrian in Arrojos Formation, in the state of Sonora, Mexico, and Chancelloria sp. from the early Cambrian in Puerto Blanco Formation, in association with ?Kiwetinokia in Cerro Rajon in Caborca region, being this the first record for San Jose de Gracia, central Sonora.
Keywords: Chancelorids, Protospongids, Cambrian, Sonora, Mexico.
1. Introducción
Los chancelóridos son un grupo problemático, estratigráficamente restringidos al Cámbrico. Estos ocurren desarticulados y asociados a ricas shelly faunas. Los chancelóridos incluyen individuos con cuerpos en forma de bolsa (escleritomo) y cubiertos con espinas comúnmente compuestas llamadas escleritos.
Los escleritomos de chancelóridos enteramente preservados son raros, mientras que los escleritos disociados están ampliamente distribuidos en rocas especialmente del Cámbrico medio. Hacia el Cámbrico tardío el registro muestra una rápida declinación por lo que se interpreta que probablemente se extinguieron antes de finales del Cámbrico.
Especímenes casi completos de Chancelloria son conocidos de Burgess Shale en Canadá (Walcott, 1920), de la Formación Wheeler, EUA (Rigby, 1978) y de la Formación Kalili, Guiizhou, China (Zhao et al., 1999).
Los primeros escleritos y fragmentos de chancelóridos fueron descritos por primera vez por Walcott (1920) de la colección del yacimiento de Burgess Shale del Cámbrico medio, en la Columbia Británica, quien los incluyó en la Clase Heteractinellida, dentro de los Porifera (Rigby, 1978; Janussen et al., 2002). Sin embargo, investigaciones recientes sugieren que el modo de crecimiento de los escleritos de este organismo es diferente al encontrado en las esponjas, considerándolo un organismo problemático del Cámbrico que produce escleritos similares a los de los Porifera (Elliott, 1987). Goryansky (1973) notó que los escleritos consistían en un compuesto de carbono-fosfato por lo que sugirió que serían estructuras externas más que espículas. Bengtson y Missarzhevsky (1981) los asignaron al grupo extinguido Coeloscleritophora (animales que poseen epitelio) para incluir metazoos como Wiwaxiidae Walcott, 1911; Siphogonuchitidae Qian, 1977 y Chancelloridae Walcott, 1920 dentro de un phylum indeterminado (Janussen et al., 2002; Elliott, 1987). Ax (1995) crea el grupo epitheliozoa un clado (no un phylum) que incluye a todos los eumetazoa y homoscleromorpha.
Sperling et al., 2007, argumentan que el proceso de “espiculación” es convergente con Porifera y en consecuencia los chancelóridos podrían pertenecer a este grupo de organismos bentónicos, aun teniendo en cuenta las claras diferencias de biomineralización entre las espículas y los escleritos.
La Familia Chancelloriidae actualmente comprende 13 géneros en parte controversiales, siendo los más comunes Chancelloria Walcott, 1920, Allonia Doré y Reid, 1965 y Archiasterella Sdzuy, 1969. La mayoría de los ejemplares de Chancelloria para América del sur se conocen solo por sus escleritos, ya que no han sido encontrados ejemplares completos (Beresi y Rigby, 1994).
Las esponjas (phylum Porifera) están entre los metazoos más primitivos y habitan los océanos desde el Precámbrico Superior. Estos organismos secretan una variedad de esqueletos minerales (espículas) de calcita, aragonita y/o sílice amorfa que confiere resistencia y las protege de las perturbaciones físicas.
La forma de la espícula es controlada genéticamente por un filamento axial orgánico (proteína), sin embargo, experimentos recientes han demostrado la influencia de los factores ambientales en la modulación del tamaño, forma y la presencia o ausencia de uno o más tipos de espículas. Las consecuencias de dichas variaciones pueden ser importantes en la taxonomía de Porifera (Uriz et al., 2003).
Las espículas de las hexactinéllidas se forman en un ambiente que es subsaturado con respecto a la sílice. La silicificación es el proceso dominante de biomineralización en esponjas silíceas de las clases Hexactinéllida y Demospongiae. El tipo básico simple de espícula en las hexactinélidas es la hexactina que puede fusionarse en los extremos de los rayos de las espículas adyacentes o hasta formar un esqueleto rígido.
Las muestras calcáreas del Cámbrico medio de San José de Gracia contienen escasos fragmentos del esqueleto espicular semi-articulado característico de las protoesponjas y asociaciones de espículas de la clase Hexactinellida. Las espículas son de dos categorías: estauractinas y hexactina-pentactinas de diversos tamaños y están piritizadas. Se encuentran irregularmente distribuidas sobre el plano de estratificación de calcipelitas delgadas rojizas, como resultado del transporte y redepositación.
2. Localización
El material estudiado procede del Cerro El Chihuarruita, con coordenadas geográficas 29° 17’ Latitud N y 110° 35’ Longitud W, ubicado al sureste del ejido de San José de Gracia, en la parte central del estado de Sonora. San José de Gracia es la población más cercana a Hermosillo, situada a 36 kilómetros al NW del afloramiento y es la ciudad más importante próxima al área de estudio (Figura 1).
El área de estudio presenta una historia geológica compleja y ha sido descrita por diversos autores (Stewart et al., 2002; Cuen et al., 2009; Nardin et al., 2009; Buitrón et al., 2011; Cuen, 2012) con énfasis en la estratigrafía y paleontología de las rocas cámbricas.
Las rocas más antiguas del área son del Paleozoico inferior, constituidas principalmente por caliza, lutita y arenisca; que afloran al SE del ejido de San José de Gracia, se encuentran cubiertas por rocas volcánicas terciarias.
Los primeros estudios paleontológicos formales del área fueron hechos por Nardin et al. (2009) quienes recolectaron ejemplares de Gogia granulosa Robison, 1965, así como fragmentos de hiolíthidos, trilobites y algas oncolíticas; sin embargo no incluyen dentro de sus registros a Chancelloria eros, Diagoniella sp. o taxones afines.
El Cerro El Chihuarruita forma una prominencia aislada con elevación de 500 metros. Está constituido por una potente secuencia sedimentaria marina, cuya base no aflora debido a complejos problemas estructurales, que invierten la secuencia estratigráfica, posicionando las rocas del Cámbrico inferior en el alto topográfico y las rocas del Cámbrico medio en la parte basal del cerro y el valle (Figuras 2 y 3).
3. Estratigrafía
La secuencia estratigráfica del Cerro El Chihuarruita está representada por rocas sedimentarias del Cámbrico inferior–Cámbrico medio (Figura 4), se inicia con cuarzo-arenita basal con variaciones en el tamaño de grano que no sobrepasan el milímetro, de color guinda de forma homogénea y en ocasiones de color rosa, la cual presenta óxidos de Fe y abundante fracturamiento. Es común observar estratificación laminar y cruzada.
Estas rocas están compuestas por más del 95% de granos de cuarzo, observándose en algunas ocasiones sobre crecimiento de cuarzo rellenando espacios porosos, siendo sílice el cementante principal. En la parte superior de estas rocas se presentan horizontes de arenita de grano grueso, con gran cantidad de bioturbación e icnofósiles, entre los que se reconoce a Skolithos sp., Planolites sp., Arenicolites sp., y Palaeophycus sp. (Cuen, 2012).
Estas rocas están sobreyacidas concordantemente por un paquete constituido por caliza arenosa intercalada con lutita y caliza. La caliza arenosa es de color café rojizo y presenta granos de cuarzo angulosos a subangulosos mal clasificados, en una matriz calcárea muy alterada con óxidos. Seguida por intercalaciones de lutita y caliza con fósiles de fragmentos de trilobites como Bristolia sp. cf. B. bristolensis Resser, 1928 (Nardin et al., 2009) y ?Olenellus; así como los hiolíthidos Hyolithes sonora Lochman, 1952 y Haplophrentis reesei Babcock y Robison, 1988 (Buitrón et al., 2011); probablemente corresponda a la Biozona de Bonnia-Olenellus. Posteriormente aflora un banco de algas oncolíticas asignadas a ?Girvanellai.
Este paquete subyace de manera concordante por una caliza oolítica masiva, de color gris oscuro, la cual se encuentra ligeramente dolomitizada, formando cantiles.
Finalmente aflora un potente paquete de lutita intercalada con caliza, con abundantes fósiles del Cámbrico medio representativos de la Biozona de Albertella, entre los que destacan trilobites como Elrathina antiqua, Oryctocephalites walcotti, Peronopsis bonnerensis, Bathyuriscus sp., y Pagetia sp., entre otros (Cuen, 2012).
La fauna de braquiópodos es también abundante y está constituida por Acrothele sp., Prototreta sp., Dictyonina sp., Linnarsonia sp., Lingulella sp. y otros no identificados.
En esta unidad se recolectaron en la base y en la cima, escleritos de Chancelloria eros Walcott, 1920 así como Diagoniella sp. y asociaciones de espículas hexactinélidas de la Familia Protospongiidae.
4. Paleontología y sistemática
El material descrito e ilustrado en esta publicación está depositado en la colección de referencia de la Universidad de Sonora, México bajo el número de catálogo USDG238-248.
Clase Coeloscleritophora Bengtson y Missarzhevsky, 1981
Orden Chancelloriida Walcott, 1920
Familia Chancelloriidae Walcott, 1920
Diagnosis. Ejemplares coeloscleritóforos con armadura de escleritos como espinas. Los escleritos son usualmente compuestos, consisten en rayos individuales unidos en la base, con cavidades internas y foramen separados. La sección transversal de los rayos es casi circular, excepto donde las bases se unen. El foramen es restringido (Bengtson, 1990, p.47).
Género Chancelloria Walcott, 1920
Diagnosis. Ejemplar de forma tubular, probablemente hueco, de cuerpo blando, con una armadura de escleritos compuestos, en su mayor parte, de 6 rayos en un plano, dispuestos radialmente y equidistantes, alrededor de un disco basal central, pero los rayos pueden ser de cuatro a nueve. La longitud del organismo total es de 95 mm y con los escleritos con rayos hasta 3 mm de largo y con discos basales o centrales de aproximadamente 0.6-1.0 mm de ancho. Cada rayo hueco presenta un sólo foramen sobre un lado, cerca de la unión con las facetas basales.
Chancelloria eros Walcott, 1920
Especie tipo. Chancelloria eros Walcott, 1920
Chancelloria eros Walcott, 1920; Janussen et al., 2002, p. 599, fig. 3, p. 600, fig. 4, p.602, fig. 6; Beresi y Rigby, 1994, p. 215, fig. 5-9, p. 216, fig. 6; Rigby, 1978, p.1330, fig.3; p. 1342, fig. 5.
Diagnosis. Escleritos con rayos cilíndricos que se adelgazan hacia los extremos. Los escleritos son simétricamente radiales y tienen predominantemente siete rayos (7+1) más uno vertical, algunos pueden tener cuatro a más de ocho radios laterales, parcialmente de tamaño diferente, organizados en planos horizontales, los cuales pueden distar curvadamente hacia el exterior; los radios horizontales pueden sobresalir verticalmente, pero en algunas ocasiones están ausentes (Janussen et al., 2002).
Descripción. Escleritos aislados, caracterizados por 6 a 7 rayos tangenciales en un plano lo que se asemeja a una figura estrellada y un rayo central vertical perpendicular preservado como un nodo aplanado o protuberancia. De algunos escleritos sólo se han preservado algunos rayos tangenciales desarticulados. La base o disco central desde el cual se articulan los rayos es aparentemente abultado o discoideo con un diámetro aproximado de 0.53 mm; no se observan las facetas basales de articulación de los rayos.
Los rayos se encuentran distribuidos radialmente de la siguiente forma: cuatro escleritos de la colección son de tipo (7+1), mientras que uno presenta sólo cuatro rayos fragmentados de los siete que se insinúan en el ejemplar mal conservado y desplazado y otro es de seis rayos (6+0). Otros escleritos algo difusos y desarticulados se observan asociados a los descritos. Los escleritos son de tamaño similares a excepción de uno más pequeño algo aplastado.
Los radios divergen del centro de manera uniforme a 60° aproximadamente uno de otro. La mayoría de los rayos están fragmentados por lo que la longitud total de los mismos se desconoce. En dos escleritos se observan los moldes de radios laterales inferiores que alcanzan una longitud de 2.10, 1.98 y 1.78 mm desde la base de inserción y se curvan hacia el exterior. La base de los rayos en la articulación varía entre 0.35, 0.40 y 0.50 mm y progresivamente se adelgazan hacia el extremo terminando en punta aguzada. En general los rayos aparentan tener una superficie lisa y algo bulbosa, con un relieve que destaca sobre el plano de la roca. El nodo central que corresponde a la parte basal del rayo vertical proximal no preservado tiene un diámetro de 0.5 mm. Además de los escleritos descritos, se observan moldes de escleritos mal conservados y rayos curvados desarticulados.
Todos los escleritos de la colección están apoyados con su lado inferior sobre la superficie de la roca por lo que no es posible la observación del foramen, abertura individual de cada rayo.
Discusión. Los escleritos de San José de Gracia presentan una distribución de los rayos (7+1), longitud máxima de rayos de hasta 2.05 mm, base de los rayos de aproximadamente de 0.35mm y un disco central de 0.5-0.6 mm de diámetro. Estas características son similares a otros ejemplares de Chancelloria eros, en particular la composición de (7+1) escleritos, típica de la especie (por ej. Goryansky, 1973; Mehl, 1991). Una característica de la especie Chancelloria eros (Walcott, 1920) es que los rayos se adelgazan en las facetas de articulación con el disco central, si bien esta característica no se observa claramente en los escleritos de Sonora debido posiblemente a la preservación de los mismos, la morfología general de los escleritos, la distribución de los rayos (7+1) y el diámetro del disco central permiten asignar estos ejemplares a la especie C. eros descrita por Walcott, 1920 para la localidad de Burgess Shale. La mayoría de los ejemplares de Chancelloria para Sudamérica se conocen solo por sus escleritos, ya que no han sido encontrados ejemplares completos (Beresi y Rigby, 1994).
Material examinado. Cinco escleritos aislados. El material se encuentra depositado en la colección de referencia del Departamento de Geología de la Universidad de Sonora, México, con número de catálogo USDG 238-242.
Distribución. Cámbrico medio.
Allonnia? sp.
Descripción. Dos escleritos con estructura 2+0. Uno de los dos rayos es grueso y recurvado en un plano medio hacia el ápice en ambos escleritos y el otro rayo es recto y aparentemente en un plano horizontal. Estos rayos gruesos y recurvados tienen un diámetro basal de 0.12 mm, y 2.25 mm de longitud, mientras que los otros rayos miden 1.10 mm y 0.75 mm. Ambos escleritos parecen estar en posiciones paralelas.
Discusión. Debido a que el material se limita a dos escleritos, la morfología de ambos con dos rayos curvos permite asignar con dudas ambos escleritos al género Allonnia. Sin embargo, el esclerito de la Figura 6a parece asemejarse a Archiasterella fletchergryllus Randell et al. (2005), por su rayo central recurvado. No obstante, teniendo en cuenta la variación de la forma de escleritos en la especie y particularmente dentro de los organismos individuales y que hasta el momento A. fletchergryllus sólo se conoce del Cámbrico temprano (Branchiano) de los Territorios del noroeste de Canadá, se prefirió asignarlos con dudas al género Allonnia.
Phylum Porifera Grant, 1836
Clase Hexactinellida Schmidt, 1870
Orden Reticulosa. Reid, 1958
Familia Protospongiidae Hinde, 1887
Diagnosis. Esponjas pequeñas de forma esferoidal a vasiforme; red esqueletal de una sola capa o múltiples capas delgadas de estauractinas o pentactinas, junto con posibles rhabdodiactinas en algunas especies; espículas típicamente en arreglo paralelo, las más largas comúnmente en arreglo quincuncial (pero algunas veces con solapamiento de los rayos formando mallas cuadradas). Espículas pequeñas de varios órdenes de tamaño subdividen las mallas sin solapar ni formar arreglos cuadrados. Prostalia (posibles rhabdodiactinas) puede estar desarrollada alrededor del ósculo y proyectarse en todo el contorno. Basalia a menudo desarrollada.
Distribución estratigráfica. Cámbrico inferior–Jurásico.
Descripción. Las muestras de calcipelitas contienen asociaciones de espículas triaxiales de tipo estauractina y hexactina-pentactina, de diversos tamaños, disociadas, desarticuladas del cuerpo de la esponja, agrupadas en forma irregular sin una orientación preferencial, en algunos casos superpuestas y escasos fragmentos del esqueleto de protoesponjas. Las espículas están reemplazadas por pirita. La mayoría de estas espículas son estauractinas de tipo (+), compuestas por cuatro rayos en un plano y dos ejes que se insertan en ángulo recto de 90° y en menor proporción en ángulos menores a 45º (tipo x). La estructura espicular consiste en mallas ortogonales y romboidales (diagonales) en arreglo paralelo algo distorsionadas por aplastamiento y superposición de las espículas y en general mal conservada en parches de las muestras. Se ha observado que los rayos de las estauractinas de mayor tamaño se fusionan en los extremos (arreglo quincuncial) formando uno de los lados del cuadrángulo de la estructura esqueletal (checar figura 6b-g). Entre estas mallas se han medido dos rayos fusionados que miden entre 6 y 6.5 mm entre los centros de dos espículas adyacentes. Las estauractinas muestran rayos rectos y lisos, de forma acicular con terminación aguda y algunas pocas tienen dos rayos levemente curvados. En general los extremos de los rayos están fragmentados. Los rayos de las espículas de mayor tamaño tienen longitudes que varían entre 4.5, 3.8, 3, 2.5 a 1.2 mm, pero mayoritariamente tienen entre 2.5, 2.8 y 3 mm y entre 0.25 y 0.15 mm de diámetro basal promedio. Las espículas de menor tamaño presentan rayos de aproximadamente 0.75, 0.45, 0.35 mm de largo promedio y 0.02 mm de diámetro promedio. En la asociación se observan espículas con rayos robustos de igual longitud y nodo central. Estas espículas se interpretan como pentactinas (5 rayos) posiblemente de la capa dermal con los cuatro rayos apoyados en la pared de la esponja y el quinto hacia el exterior si bien pudieran ser hexactinas (6 rayos), con el sexto rayo directamente hacia el interior de la pared corporal. Los 4 rayos tangenciales miden 0.70 - 0.65 mm de longitud y el nodo central correspondiente al rayo vertical proximal-distal no preservado es de 0.25 a 0.35 mm de ancho. Algunas espículas largas muestran solo dos rayos tangenciales curvados con longitudes entre 1.5, 2.6 mm hasta 4.9 mm pudiendo corresponder a espículas triaxonas de la superficie dermal.
Algunos rayos largos y muy delgados (2 a 3 mm de longitud y 0.15 mm de diámetro) de tipo monaxial aparecen con un cierto grado de paralelismo, sobre relictos de estructura espicular romboidal y espículas irregularmente distribuidas. Estos rayos (Figura 6b) podrían corresponder a espículas basales de sostén, de anclaje al sustrato (root tufts) desarticuladas de la base de la esponja que no está conservada.
Material examinado. Tres calcipelitas con número variable de espículas. Número de catálogo USDG 243-245.
Discusión. La definición de la Familia Protospongidae (Finks y Rigby, 2004) incluye un amplio rango de esponjas reticuladas de pared delgada con estructura regular a subregular de mallas cuadrangulares ortogonales o romboidales. La familia como normalmente ha sido definida resulta ser ciertamente parafilética e incluye diversos grupos que podrían ser razonablemente separados en familias monofiléticas. Una restricción a la familia incluye solo formas vasiformes a esferoidales con regular a subregular estructura de mallas con estauractinas y/o espículas pentactinas y una superposición mínima de los extremos de los rayos (overlapping) (Rigby, 1978, 1986; Rigby y Murphy, 1983). Rigby (1986) en su diagnóstico genérico de Protospongia destacó la regularidad de la arquitectura del esqueleto constituido por estauractinas organizadas en 6 órdenes de tamaños y sugirió que podría ser dividida sobre la base del arreglo espicular, particularmente de las espículas de primer orden. Hinde (1888) restringió el género a aquellos ejemplares cuyas espículas fueran exclusivamente estauractinas. En el caso del género Diagoniella Rauff 1894, la característica diagnóstica es el arreglo diagonal (aproximadamente a 45º) de las estauractinas respecto al eje vertical principal del cuerpo de la esponja, a diferencia del género Protospongia que tiene arreglo vertical.
Las muestras cámbricas de Sonora contienen parches del esqueleto con mallas rectangulares y romboidales formadas por la fusión de los extremos de los rayos de las estauractinas de mayor tamaño y asociaciones de espículas de diversos tamaños dominadas por estauractinas de tipo (+) y (x) y algunas pentactinas. Las diferencias en tamaño de las espículas observadas pueden corresponder a diferentes posiciones en el cuerpo de la esponja tales como espículas corticales, principales (formando los tractos estructurales importantes), o auxiliares que se encuentran diseminadas dentro del mesohil (la capa interna de la pared del porífero) o justo debajo de la superficie. También pueden corresponder a distintos órdenes de tamaño de espículas que constituyen la estructura del esqueleto de un mismo individuo como en ambos géneros.
Las estauractinas y las mallas cuadrangulares del esqueleto son características de los géneros Protospongia y Diagoniella. Sin embargo, la posición de la estructura espicular en relación al eje del cuerpo de la esponja no se ha preservado, y no es posible una asignación genérica.
Las asociaciones de espículas con dominancia de estauractinas desarticuladas, mezcladas y a veces superpuestas de Sonora corresponden a una tanatocenosis sin una asignación genérica cierta se asignan a la Familia Protospongiidae, infiriendo que dichas asociaciones pueden corresponder tanto a Protospongia, como a Diagoniella o a ambos géneros del Orden Reticulosa.. Espículas estauractinas disociadas como las de la colección estudiada tienen amplia distribución a nivel mundial en los mares del Cámbrico inferior y medio.
Diagoniella sp.
Descripción. La forma del cuerpo de las esponjas no está bien definida pero se puede observar un contorno más o menos ovoidal a globular por la presencia de espículas y en otras partes del mismo fragmento por el oscurecimiento de la matriz rojiza donde mallas romboidales difusas están presentes. En otro fragmento se observan parches con mallas romboidales difusas. En todos los casos los parches del esqueleto preservado está constituido por estauractinas de tipo (x) fusionadas por sus extremos formando una red romboidal con ángulos entre 35º a 45º. La longitud de las espículas varía entre 2 mm y 4.0 mm. En una de las esponjas (Figura 7c), se observan dos espículas monaxonas (un solo rayo) que parecen insertarse en la base del cuerpo de la esponja como estructura de sostén (root tuft). Material examinado. Tres calcipelitas con número variable de espículas. Número de catálogo USDG 246-248.
Discusión. La estructura conservada del esqueleto está constituida por diversos fragmentos de mallas romboidales y estauractinas de tipo (x) y dos espículas monaxonas posiblemente de sostén, como en las especies Diagoniella hindei y D. coronata. Estas características permiten asignar estos ejemplares al género Diagoniella sin que el material permita la asignación específica.
Distribución. Cámbrico medio.
Protospongioideo indeterminado
Descripción. Algunas espículas escasas de la colección son triaxonas de seis rayos, hexactina-pentactinas constituidas por cuatro rayos en un plano y un nodo central en la conjunción de los dos ejes principales, correspondiente al rayo vertical proximal/distal no preservado. El diámetro del nodo de las espículas de mayor tamaño varía entre 0.40 - 0.60 mm y 0.4 - 0.5 mm de espesor preservado. Otras espículas tienen nodos centrales de 0.125 mm y rayos finos de 1.75 a 1.9 mm de longitud. Numerosos nodos redondos aislados se han preservado piritizados sobre los planos de estratificación con diámetros de 0.18, 0.14 hasta 0.10 mm en otros casos ha quedado un hueco en el plano de estratificación. En general los rayos de las espículas de mayor tamaño tienen longitudes similares de 4.20 a 2.20 mm y diámetros basales de 0.35 - 0.25 mm en promedio. Otras hexactinas de menor tamaño presentan de 0.75 y 0.65 mm de longitud y nodo central de 0.35 mm. Los rayos son lisos y rectos pero en algunas espículas dos o cuatro rayos tangenciales son ligeramente curvos y sus longitudes varían entre de 1.15, 3.75 y 4.9 mm.
Discusión. El tipo de espícula hexactinélida básica es la hexactina simple, mientras que otras triaxonas tales como pentactinas y estauractinas con cinco o cuatro rayos respectivamente, son formas derivadas, simplificadas por reducción de rayos (Mehl, 1991). Estas espículas son muy comunes en la mayoría de los grupos de las Hexactinélidas y asociadas a simples estauractinas, son características de diferentes géneros cámbricos de la Familia Protospongiidae, como Gabelia del Cámbrico inferior-Silúrico de China, Canadá y Estados Unidos y Triticispongia diagonata del Cámbrico inferior de China, entre otros. La falta de preservación del esqueleto articulado en las muestras analizadas no permite una asignación genérica.
5. Conclusiones
Se describen las especies Chancelloria eros Walcott, 1920, Diagoniella sp., Allonnia? sp. y asociaciones de espículas hexactinélidas estauractinas y hexactina-pentactinas asignadas a la Familia Protospongiidae Walcott 1920, procedentes de afloramientos del Cámbrico medio de la región de San José de Gracia, Sonora central. La asociación biótica está constituida por Peronopsis bonnerensis Resser, 1938, Oryctocephalites walcotti Reeser, 1938, Elrathina antiqua Palmer y Halley, 1979 y Ogygopsis typicalis Resser, 1939.
La presencia de chancelóridos y de protoesponjas hexactinélidas corrobora la abundancia y distribución global de estos metazoos en mares someros, cálidos del periodo Cámbrico.
Agradecimientos
Los autores agradecen al Dr. Guadalupe Inocente Espinoza, Jefe del Departamento de Geología de la Universidad de Sonora y a la Dra. Elena Centeno García, Directora del Instituto de Geología de la UNAM por el apoyo brindado para la realización de esta investigación. F. Cuen agradece las facilidades otorgadas para llevar a cabo la investigación al Ing. Miguel Ángel Soto y Bedolla de la Dirección de Exploración de Timmins Gold Corp. México S.A. de C.V.
Se reconoce a los Dres. Rogelio Monreal Saavedra, del Departamento de Geología de la Universidad de Sonora, Francisco J. Vega Vera, editor asociado y Juan Francisco Sánchez Beristaín, del Instituto de Geología y de la Facultad de Ciencias, UNAM, respectivamente, el haber revisado el manuscrito y hacer importantes observaciones. Al maestro José Carlos Jiménez López del posgrado en Ciencias de la Tierra del Instituto de Geología, UNAM, quien colaboró en la elaboración de las figuras.
Asimismo, se agradece el apoyo económico proporcionado por los proyectos UNAM-DGAPA-PAPIIT No. IN105012 y CONACYT No. 165826 para la realización del trabajo de campo y estancias académicas.
Referencias
Ax, P., 1995, Das System der Metazoa 1, Ein Lehrbuch der phylogenetischen Systematik. Gustav Fischer Verlag, Stuttgart, 226 p.
Babcock, E.L., Robison, A.R., 1988, Taxonomy and paleobiology of some Middle Cambrian Scenella (cnidaria) and Hyolithids (Mollusca) from Western North America: Paleontological Contributions, The University of Kansas Press, 121, 1-22.
Bengston, S., 1990, Chancelloriids, en Bengston, S., Conway Morris, S., Cooper, B.J., Jell, P.A., Runnegar, B.N. (eds.), Early Cambrian fossils from South Australia: Association of Australasian Palaeontologists, Memory 9, 45-69.
Bengtson, S., Missarzhevsky, V., 1981, Coeloscleritophora - a major group of enigmatic Cambrian metazoans: United States Geological Survey, Open-File Report 81-743, 19-21.
Beresi, M.S., Rigby, K.J., 1994, Sponges and Chancelloriids from the Cambrian of Western Argentina: Journal of Paleontology, 68, 208-217.
Buitrón, B.E., Vachard, D., Clausen, S., Gómez, E.C., 2011, Cambrian mollusks (Hyolithida, Syssoiev, 1957) from San José de Gracia, Sonora, Northwestern Mexico: Reunión Internacional de Malacología, Libro de resúmenes, 175 p.
Cuen, R.F., Almazán, E., Montijo, A., Minjarez, I., Grijalva, F., Monreal, R., Schwenicke, T., Ochoa, J., 2009, Faunas marinas de invertebrados del Cámbrico temprano y medio en la parte central del Estado de Sonora, México. Geos, Unión Geofísica Mexicana. Libro de resúmenes, 29, 91 p.
Cuen, R.F., 2012, Sobre la Estratigrafía y Paleontología del Cámbrico del área de San José de Gracia, Sonora central, México, Universidad de Sonora, Departamento de Geología, tesis de Licenciatura, 65 p.
Doré, E., Reid, R.E.H., 1965, Allonnia tripodophora nov. gen., nov. sp., nouvelle éponge du Cambrien inférieur de Carteret (Manche): Comptes Rendus Sommaires Séances Société Géologique de la France, 20-21.
Elliott, D.K., Martin, D.L., 1987, Chancelloria, an Enigmatic fossil from the Bright Angel Shale (Cambrian) of Grand Canyon, Arizona: Journal of the Arizona-Nevada Academy of Science, 1, 67-72.
Finks, R.M., Rigby, J.K., 2004, Paleozoic Demosponges, en Kaesler, R.L. (ed.), Treatise on Invertebrate Paleontology, Part E, Porifera Revised, 3. Geological Society of America and the University of Kansas Press, E1-E173.
Goryansky, V.Y., 1973, On the necessity of excluding the genus Chancelloria Walcott from the phylum Porifera: Trudy Institut Geologii i Geofiziki So AN SSSR 49, 3944. (in Russian).
Grant, R.E., 1836, Animal Kingdom, en Todd, R.B. (ed.), The Encyclopaedia of Anatomy and Physiology, Sherwood, Gilbert & Piper, London, 1, 107-118.
Hinde, G.J., 1887-1888, A monograph of the British Fossil sponges: Palaeontographical Society of London, 1-92 (1887); 93-188 (1988).
Janussen, D., Steiner, M., Zhu, M., 2002, New well-preserved scleritomes of Chancelloridae from the Early Cambrian Yuanshan Formation (Chengjiang, China) and the Middle Cambrian Wheeler Shale (Utah, USA) and Paleobiological Implications: Journal of Paleontology, 76, 596-606.
Mehl, D., 1991, Are Protospongiidae the stem group of modern Hexactinellida?, en Reitner J., Keupp, H. (eds.), Fossil and recent sponges, Berlin, 44-53.
Nardin, E., Almazán, E., Buitrón, B.E., 2009, First report of Gogia (Eocrinoidea, Echinodermata) from the Early-Middle Cambrian of Sonora (Mexico), with biostratigraphical and palaecological comments: Geobios, 42, 233-242.
Palmer, A.R., Halley, R.B., 1979, Physical Stratigraphy and Trilobite Biostratigraphy of the Carrara Formation (Lower and Middle Cambrian) in the Southern Great Basin: United States Geological Survey Professional Paper 1047, 1-139.
Qian, Y., 1977, Hyolitha and some problematica from the Lower Cambrian Meishucun Stage in Central and SW China: Acta Palaeontologica Sinica, 16, 255-278.
Randell, R.D., Lieberman, B.S., Hasiotis, S.T., Pope, M.C., 2005, New chancelloriids from the Early Cambrian Sekwi Formation with a comment on chancelloriid affinities: Journal of Paleontology, 79, 987–996.
Rauff, H., 1894, Palaeospongiologie: Palaeontographica, 40, 233-346.
Reid, R.E.H., 1958, A monograph of the Upper Cretaceous Hexactinellida of Great Britain and Northern Ireland, part I: Palaeontographical Society Monograph, London, i-xlvi.
Resser, C.E., 1928, Cambrian fossils from the Mohave Desert: Smithsonian Miscellaneous Collections, 81, 1-14.
Resser, C.E., 1938, Middle Cambrian fossils from Pend Oreille Lake, Idaho: Smithsonian Miscelaneous Collection, 97, 1-12.
Resser, C.E., 1939, The Spence Shale and its fauna: Smithsonian Miscellaneous Collections, 12, 1-29.
Rigby, K.J., 1978, Porifera of the Middle Cambrian Wheeler Shale, from the Wheeler Amphitheater, House Range, in Western Utah: Journal of Paleontology, 52, 1325-1345.
Rigby, K.J., 1986, Sponges of the Burgess Shale (Middle Cambrian British Columbia): Palaeontographica Canadiana, Monograph 2, 1-105.
Rigby, K.J., Murphy, M.A., 1983, Gabelia, a new Late Devonian lyssakid protosponge from the Roberts Mountains, Nevada: Journal of Paleontology, 57, 797–803.
Schmidt, O., 1870, Grundzuge einer Spongien-Fauna des atlantischen Gebietes, Jena, Leipzig, iv +88.
Sperling, E.A., Pisani, D., Peterson, K.J., 2007, Poriferan paraphyly and its implications for Precambrian palaeobiology, en Vickers-Rich, P., Komarower, P. (eds.), The Rise and Fall of the Ediacaran Biota: Geological Society, London, Special Publications 286, 355–368.
Stewart, J.H., Poole, F.G., 2002, Inventory of Neoproterozoic and Paleozoic Strata in Sonora, Mexico. United States Department of the Interior and United States Geological Survey. Open-File Report 902-97, 50 p.
Uriz, M.J., Turon, X., Becerro, M.A., Agell, G., 2003, Siliceous spicules and skeletons frameworks in sponges: origin, diversity, ultrastructural patterns, and biological functions: Microscope Research and Technique, 62, 279-99.
Walcott, C.D., 1911, Cambrian geology and paleontology, Middle Cambrian annelids: Smithsonian Miscellaneous Collections, 57, 109-145.
Walcott, C.D., 1920, Middle Cambrian Spongiae: Smithsonian Miscellaneous Collections 67, 261-364.
Zhao, Y.L., Yuan, J.L., Zhu, M.Y., Yang, R.D., Qian, Y., Huang, Y.Z., Guo, Q.J., Pan, Y., 1999, A progress report on research on the early Middle Cambrian Kaili biota, Guizhou Province, PRC: Acta Palaeontologica Sinica, 38, (in Chinese with English abstract), supplement, 1-14.
Manuscrito recibido: Enero 8, 2013.
Manuscrito corregido recibido: Marzo 7, 2013.
Manuscrito aceptado: Abril 6, 2013.
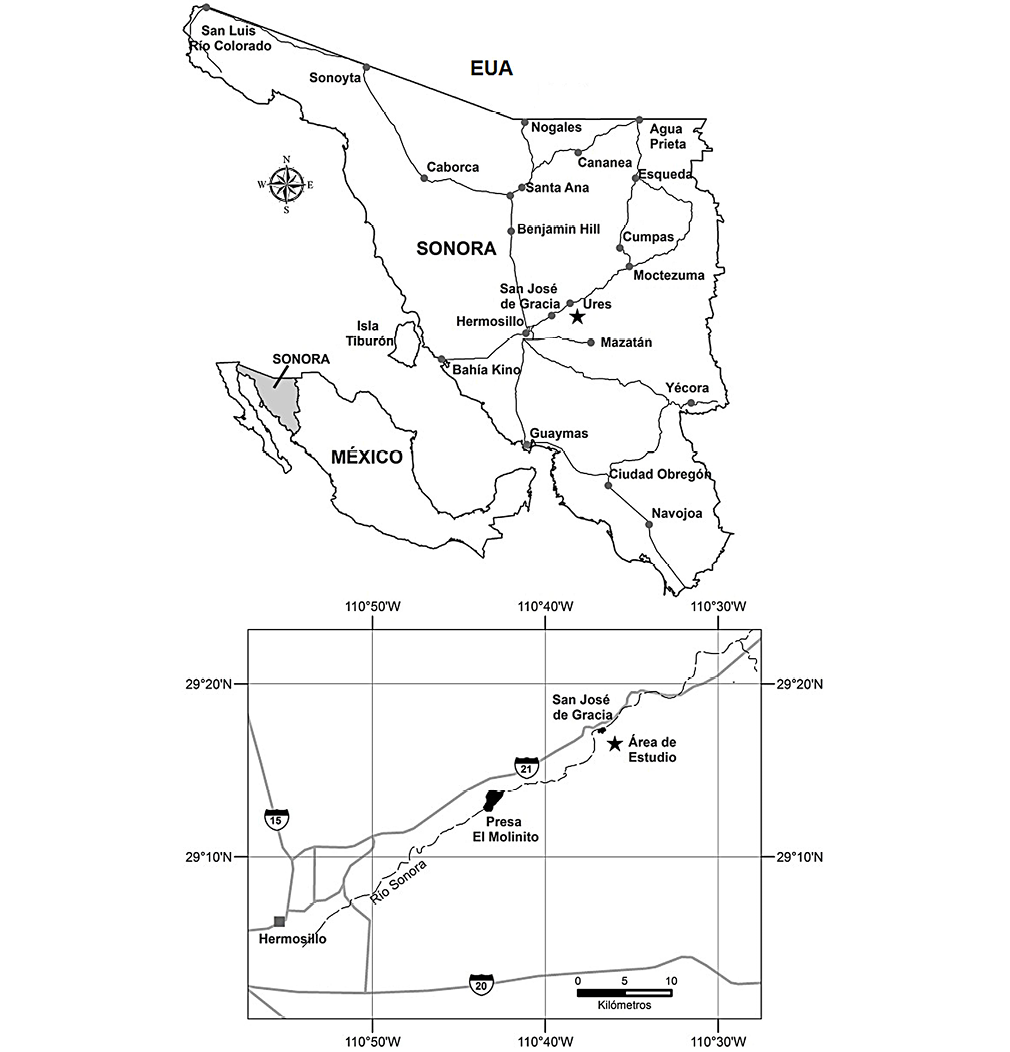
Figura 1. Localización del área de San José de Gracia, Sonora, (Cuen, 2012).

Figura 2. Fotografía panorámica del Cerro El Chihuarruita, vista hacia el noreste (Cuen, 2012).
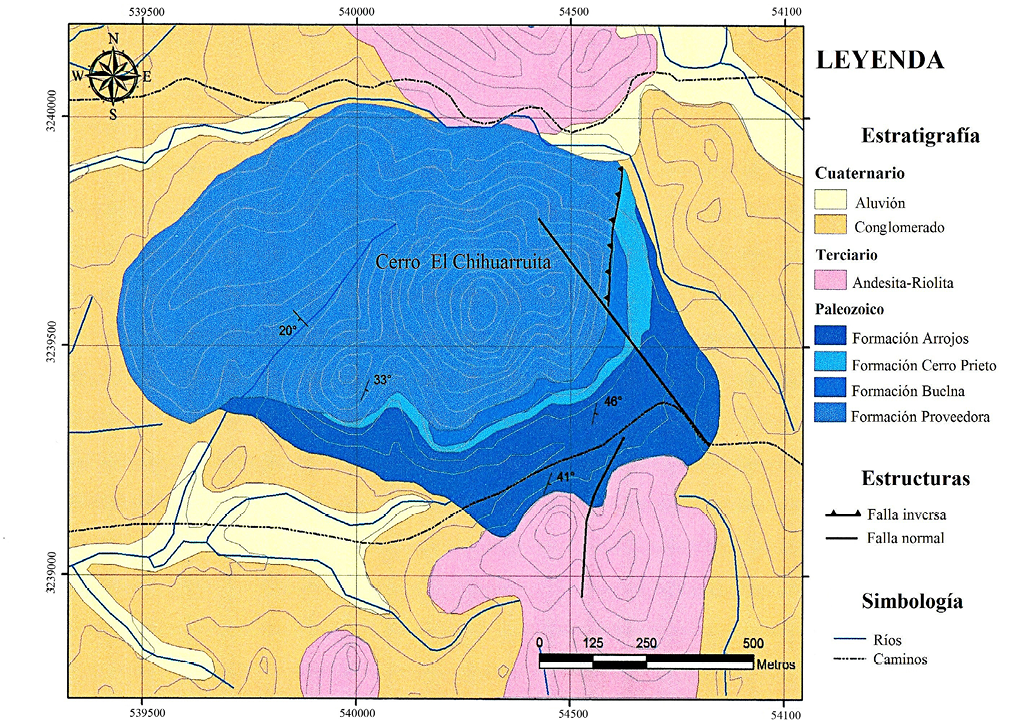
Figura 3. Mapa Geológico del Cerro El Chihuarruita San José de Gracia, Sonora (Cuen, 2012).
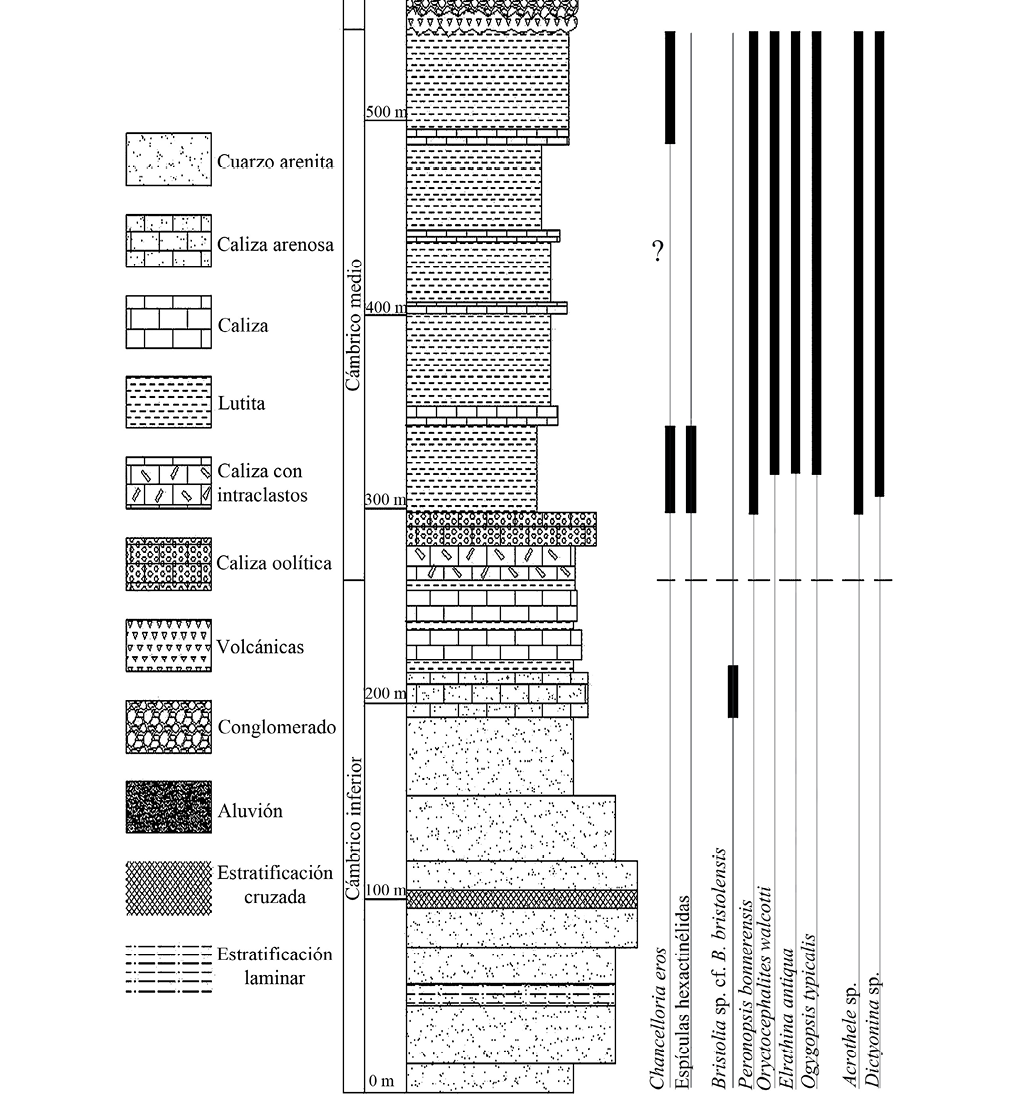
Figura 4. Columna estratigráfica del Cerro El Chihuarruita en San José de Gracia, Sonora (Cuen, 2012).

Figura 5. Chancelloria eros Walcott, 1920. a-d. Escleritos con rayos cilíndricos que se adelgazan hacia los extremos (Cuen, 2012).

Figura 6. Allonnia? sp. a, b. Escleritos de Allonnia? sp. y estauractinas dispersas; c. Acrothele sp. y posible esclerito de Allonnia sp. sobre relicto de malla espicular; d. Arreglo quincuncial de grandes estauractinas con los extremos de los rayos fusionados (flecha); e. Estauractinas con rayos unidos forman un cuadrilátero; uno de los rayos es curvo; f. Estauractinas tipo (+) con rayos fusionados y cranidio de trilobite; g. Estauractinas con rayos curvos posiblemente correspondan al margen de una esponja sobre esqueleto espicular difuso y grandes estauractinas, y cranidio de trilobite (Elrathina antiqua Palmer) y estauractinas difusas, (Cuen, 2012).
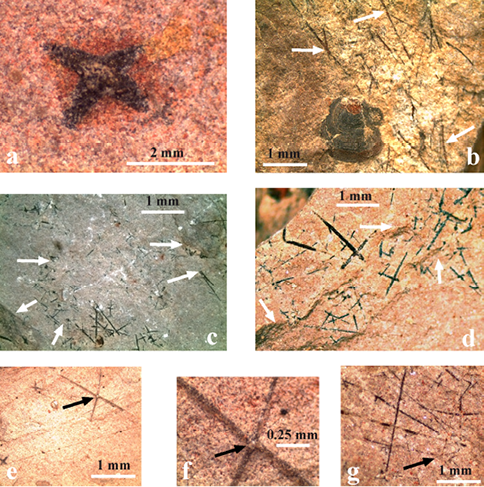
Figura 7. Diagoniella sp. a. Hexactina-pentactina. El nodo central corresponde al rayo vertical proximal-distal no preservado; b. Fragmentos de la arquitectura espicular romboidal posiblemente de Diagoniella sp. Largos rayos rectos podrían corresponder a espículas de sostén (flechas); c. posible esponja globular con relictos de esqueleto (centro) y en margen inferior izquierdo (señaladas con flechas); d. estructura esqueletal romboidal en margen inferior izquierdo y centro, estauractinas de tipo (+) desarticuladas de diversos tamaños; e, f. Hexactina-pentactina. La flecha indica el nodo central correspondiente al rayo vertical proximal-distal no preservado (figura 6f X3); g. Grandes estauractinas sobre relictos de estructura esqueletal (flecha), (Cuen, 2012).