
|
BOLETÍN DE LA SOCIEDAD GEOLÓGICA MEXICANA Vol 65, Núm. 3, 2013, P. 573-580 http://dx.doi.org/10.18268/BSGM2013v65n3a10 |
 |
Variación de la dieta del mamut de las praderas de Laguna de las Cruces, San Luis Potosí, México
Víctor Adrián Pérez-Crespo1,2,*, Joaquín Arroyo-Cabrales2, Mouloud Benammi3, Oscar J. Polaco2, Antonio Santos-Moreno4, Pedro Morales-Puente5, Edith Cienfuegos-Alvarado5, Francisco J. Otero5
1Posgrado en Ciencias, CIIDIR-Oaxaca-IPN, Calle Hornos 1003, Sta. Cruz Xoxocotlán, Oaxaca.
2Laboratorio de Arqueozoología "M.en C. Ticul Álvarez Solórzano", Subdirección de Laboratorios y Apoyo Académico, Instituto Nacional de Antropología e Historia, Moneda 16, Col. Centro, Ciudad de México 06060.
3Institut International de Paléoprimatologie Humaine: Evolution et Paléoenvironnements (IPHEP)-UMR CNRS 6046, SFA-Université de Poitiers, Bât. de Sciences Naturelles (3ème étage). 40 avenue du Recteur Pineau, F86022 Poitiers Cedex, Francia.
4Laboratorio de Ecología Animal, CIIDIR-Oaxaca-IPN, Calle Hornos 1003, Sta. Cruz Xoxocotlán, Oaxaca.
5Instituto de Geología, Universidad Nacional Autónoma de México, Circuito de la Investigación Científica S/N, Ciudad Universitaria, Coyoacán, Ciudad de México, 04150.
* This email address is being protected from spambots. You need JavaScript enabled to view it.
Resumen
Usando técnicas biogeoquímicas basadas en la cuantificación de 13C/12C y 18O/16O presentes en el esmalte dental, se infirió si la dieta de cuatro individuos del mamut de las praderas (Mammuthus columbi) de Laguna de las Cruces (Salinas, San Luis Potosí, México) cambiaba o se mantenía en el tiempo, en respuesta a las variaciones de la temperatura. Los resultados indican que la dieta del individuo DP-1975 se mantuvo sin cambio, mientras que en los mamuts DP-1976 y DP-1978 hubo una ligera tendencia a consumir más plantas C3 que C4, posiblemente debido a un descenso en la temperatura, tal como lo muestran los valores de δ18O. En contraste, los valores isotópicos de carbono del individuo DP-1979 muestran que éste poseía una dieta mixta C3/C4 que luego modificó a una exclusivamente basada en plantas C4, debido quizás a un aumento en la temperatura. Asimismo, las comparaciones efectuadas con los valores de δ13C y δ18O a nivel poblacional indican que los dos individuos jóvenes con edades de 20 y 18 años son diferentes a los individuos adultos que exhiben edades de 43 y 47 años respectivamente. Es posible que estas diferencias se deban a la conducta alimentaria diferente de los individuos de edades distintas, aunque también esto podría indicar que estos individuos provenían de diferentes áreas.
Palabras clave: Mammuthus columbi, dieta, hábitat, Laguna de las Cruces, México.
Abstract
Using biogeochemical techniques based on 13C/12C and 18O/16O relationships in dental enamel, it was inferred if the diets of four Columbian mammoths (Mammuthus columbi) from Laguna de las Cruces (Salinas, San Luis Potosí, México) changed or were similar in time as a response to temperature variations. The results show that the diet of individual DP-1975 kept unchanged, whereas for mammoths DP-1976 and DP-1978 there was a small tendency for eating more C3 plants than C4 plants, possibly due to decreased temperatures, as shown by δ18O values. However, the isotopic carbon values of mammoth DP-1979 show that the specimen was first a mixed feeder C3/C4 and later ate mostly C4 plants, probably due to an increase in the temperature. Furthermore comparisons made between δ13C and δ18O values at the population level show that the two young mammoths, with 18 and 20 years old, are different from the two adults, which seem to be 43 and 47 years old respectively. Such differences may be due to different feed behaviors among young and adult, although it could also indicate that the individuals came from different areas.
Keywords: Mammuthus columbi, diet, habitat, Laguna de las Cruces, México.
1. Introducción
Uno de los objetivos de los estudios paleoecológicos es documentar los cambios climáticos que hubo en el pasado y asociar los cambios evolutivos a estos cambios en el clima, usando diversas metodologías como los estudios morfofuncionales, análisis taxonómicos y uso de marcadores biogeoquímicos (Andrews, 1996).
En México, este tipo de estudios enfocados a los mamíferos del Pleistoceno Tardío han tratado de comprender cómo las fluctuaciones climáticas que existieron en este periodo afectaron la composición, la distribución e incluso provocaron la extinción de varias especies (Montellano-Ballesteros y Jiménez-Hidalgo, 2006). Sin embargo, la mayor parte de estos trabajos se han realizado desde un enfoque taxonómico, comparando los restos óseos de los mamíferos pleistocénicos con aquéllos que están presentes en sus contrapartes actuales, y sólo en pocos se han usado aproximaciones morfofuncionales (Bravo-Cuevas et al., 2011) y biogeoquímicas (Pérez-Crespo et al., 2009, 2010), lo cual no permite detectar si existen cambios en la dieta de un individuo o poblaciones enteras como respuesta a cambios climáticos a través del tiempo.
En México, los restos del mamut de las praderas (Mammuthus columbi) se encuentran en gran número de localidades pleistocénicas (Arroyo-Cabrales et al., 2007), una de estas localidades es Laguna de las Cruces, en el estado de San Luis Potosí, donde aparte de esta especie se han encontrado restos de bisontes (Bison bison), camellos (Camelops hesternus) y caballos (Equus sp.). Los análisis isotópicos realizados en caballos, camellos y mamuts han revelado que estas especies tenían una alimentación del tipo mixta C3/C4 y habitaban en un bosque abierto durante el Pleistoceno Tardío (Pérez-Crespo et al., 2012). Sin embargo, debe considerarse que la dieta de estos animales en forma individual pudo modificarse en el transcurso del tiempo debido a cambios en el clima o debido a que los animales se movían de un sitio a otro. En el caso de los mamuts, estos cambios quedan reflejados durante el periodo de formación de los molares, por lo que se pueden inferir variaciones en el clima de un sitio pero además, si la dieta de un mamut se modifica debido a tales fluctuaciones (Metcalfe y Longstaffe, 2012). Por lo tanto, el propósito de este artículo es analizar si la dieta de la población de mamuts de este sitio se mantuvo constante en el tiempo o si ésta se modificaba, usando para ello la composición isotópica de carbono y oxígeno presente en el esmalte.
1.1. Isótopos de carbón y oxígeno
Las relaciones de 13C/12C que se pueden analizar en el apatito del esmalte dental han mostrado ser especialmente útiles para inferir la dieta de mamíferos extintos (Ericson et al., 1981; Koch et al., 1994; MacFadden et al., 1994; Koch, 1998; MacFadden et al., 1999), al permitir comprobar las hipótesis previas obtenidas de datos morfológicos (MacFadden et al., 1994). El carbono ingresa a los animales por medio de las plantas que consumen los herbívoros y está relacionado con la vía fotosintética por la cual fue fijado (Koch et al., 1991). La forma C3 es la vía fotosintética más antigua de las plantas y el primer producto obtenido en el proceso es una molécula de tres carbonos, la ribulosa 1,5 bifosfato carboxilasa/oxigenasa (rubisco); las plantas que presentan esta vía son árboles y arbustos dicotiledóneos y algunas herbáceas de zonas frías (Ehleringer et al., 1997) y muestran valores de δ13C que van de -22 ‰ a -30 ‰ y un promedio de -27.1 ± 2.0 ‰ (Smith y Epstein, 1971; Van der Merwe y Medina, 1989; Medrano y Flexas, 2000).
En contraste, en la vía fotosintética C4, presente en herbáceas monocotiledóneas y algunos árboles y arbustos de zonas tropicales y áridas, el primer producto obtenido es una molécula de cuatro carbonos, la fosfoenolpiruvato carboxilasa (PEPC) y su δ13C promedio es de -13.1 ± 1.2 ‰, con un intervalo de valores que va de -10 ‰ a -14 ‰ (O’Leary, 1981; Cerling et al., 1997; Dawson et al., 2002; Keeley y Rundel, 2003). Una tercera vía, la CAM (metabolismo ácido crasuláceo) no puede ser distinguida isotópicamente ya que sus valores de δ13C abarcan a los de las plantas C3 y C4 (Andrade et al., 2007).
Al ser consumidas las plantas por los herbívoros, éstos tendrán valores de δ13C similares al de las plantas que se alimentaron; sin embargo los valores van a estar enriquecidos en 14 ‰ (DeNiro y Epstein, 1978; Cerling y Harris, 1999). Esto significa que los valores de δ13C de los herbívoros que se alimenten con plantas C4 (pacedores) son de -2 ‰ a +2 ‰, de -9 ‰ a -19 ‰ para los animales ramoneadores que consuman plantas C3 y los organismos que consuman ambos tipos de plantas tendrán valores intermedios de -2 ‰ a -9 ‰ (Hofmann y Stewart, 1972; MacFadden y Cerling, 1996).
En cambio, la composición isotópica del oxígeno se encuentra influida principalmente por el agua que se bebe y por el balance que existe entre el oxígeno que se pierde a través de la orina, el sudor, las heces y la exhalación de vapor de agua y CO2 y, el oxígeno que ingresa al cuerpo a través la respiración y, en menor grado, del agua de los alimentos (Koch et al., 1994). Factores como la fisiología, la dieta y el tipo de hábitat de cada especie, así como el clima, intervienen en este balance, aunque en el caso de los mamíferos de talla grande depende principalmente de este último factor (Sánchez, 2005; Koch, 2007), por lo que la composición isotópica del oxígeno del apatito puede ser utilizada para inferir las condiciones climáticas del pasado (Longinelli y Nuti, 1973; Fricke y O’Niel, 1996; Sánchez, 1997; Stuart-Williams y Schwarcz, 1997). Asimismo la información dada por este isótopo, en conjunto con los que se obtienen del δ13C, permite conocer las variaciones en la dieta de un individuo debidas a los cambios en el clima o la temperatura local (Feranec y MacFadden, 2000; Kingston y Harrison, 2007).
2. Área de estudio
Laguna de las Cruces se encuentra al oeste del poblado de Salinas en el estado de San Luis Potosí, México (22º43’N, 102º01’W) (Mirambell, 1982; Figura 1). Se realizaron excavaciones arqueológicas controladas, encontrándose restos óseos de Mammuthus columbi, Equus sp., Camelops hesternus y Bison bison, proponiéndose que dicho conjunto faunístico perteneció a la Edad de Mamíferos Norteamericana Rancholabreana (Pleistoceno Tardío) por la presencia del bisonte, acotada generalmente entre 120000 y 10500 años antes del presente (Bell et al., 2004); sin embargo, la localidad carece de fechado alguno que permita precisar una edad absoluta. La estratigrafía local de los sedimentos se encuentra representada por calizas y lutitas del Cretácico y, sobre éstas, rocas ígneas, suelos rojos, turbas y sedimentos lacustres (Figura 2), siendo en estos últimos donde fueron encontrados los restos fósiles (Flores, 1982; Reyes, 1982).
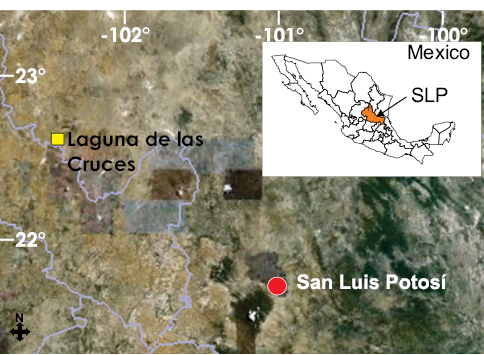
Figura 1. Localización geográfica de Laguna de las Cruces, San Luís Potosí México. Tomado de Pérez-Crespo et al. (2012).
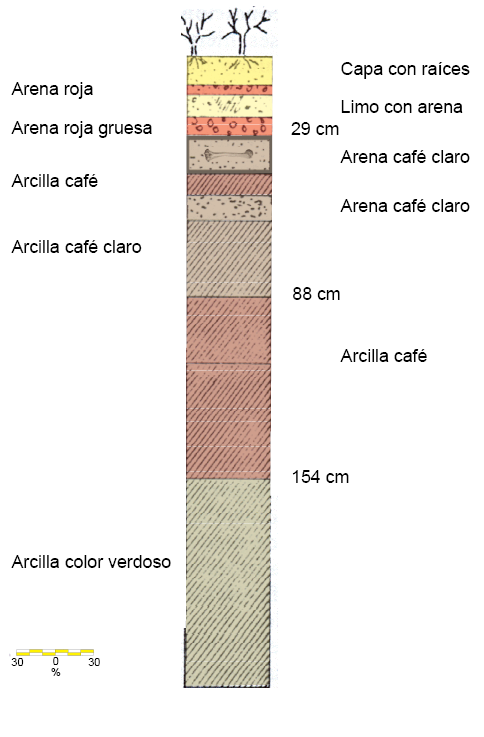
Figura 2. Estratigrafía de Laguna de las Cruces. Modificada de Flores (1982).
3. Métodos
3.1. Extracción y preparación de muestras
Los molares usados fueron encontrados en la misma capa estratigráfica que no mostraba alteración alguna, por lo que se puede proponer que son contemporáneos (Álvarez, 1982) y están bajo resguardo de la Colección Paleontológica del Laboratorio de Arqueozoología, Subdirección de Laboratorios y Apoyo Académico, Instituto Nacional de Antropología e Historia con números de catálogo DP-1975, DP-1976, DP-1978 y DP-1979.
Por medio de un taladro Dremel provisto de una fresa dental, se extrajeron series de muestras de una placa del molar DP-1979 y en dos placas del DP-1978, tomadas éstas cerca de la parte inferior de los molares (raíz) hacia la parte superior (corona); en cambio en los molares DP-1975 y DP-1976 las series de muestras fueron tomadas de la zona oclusal. Antes de comenzar a extraer el esmalte, los molares fueron limpiados con alcohol etílico y una espátula.
Una vez extraído el esmalte, éste se preparó siguiendo la metodología de Koch et al. (1997) que describimos a continuación de forma resumida: primero se pulverizó 20 miligramos de esmalte y se tamizó con malla de 125 micrómetros con objeto de obtener un polvo fino y uniforme. Se le agregaron 10 ml de H2O2 al 30 % para eliminar la materia orgánica por dos horas y después se centrífugo y se descartó el agua oxigenada; posteriormente se realizaron tres lavados a la muestra, descartando con agua destilada el agua en cada lavada. Una vez concluido el enjuague, se agregó 5 ml de una solución buffer de CaCH3CO2 - CH3COOH, 1M, pH = 4.75, dejándose reposar durante nueve horas. Posteriormente la solución buffer se descartó y las muestras volvieron a ser enjuagadas tres veces con agua destilada.
Finalmente, para eliminar el agua se les agregó alcohol etílico y se dejaron secar en un horno a 90 ºC durante 12 horas. Las determinaciones de la abundancia isotópica de las muestras se realizaron en un espectrómetro de masas Finnigan MAT 253, con sistema dual de introducción de muestras y un equipo auxiliar denominado Gas Bench con un automuestreador GC Pal con plancha de aluminio de temperatura controlada acoplado al espectrómetro de masas (Révész y Landwehr, 2002). Los resultados obtenidos fueron expresados en δ13CVPDB y δ18OVPDB y se normalizaron utilizando NBS-19, NBS-18 y LSVEC, a la escala VPDB de acuerdo con las correcciones descritas por Coplen (1988), Coplen et al. (2006) y Werner y Brand (2001). Para esta técnica se tiene una desviación estándar de 0.2 ‰ para oxígeno y 0.2 ‰ para el carbono.
3.2. Análisis estadístico
Se obtuvieron la media y la desviación estándar de los valores isotópicos de carbono y oxígeno del conjunto de muestras pertenecientes a cada individuo, así mismo, los valores individuales fueron graficados siguiendo el modelo propuesto por Feranec y MacFadden (2000) con el objeto de ver si existía un patrón que indicara cambio en la dieta, producto de las variaciones en el clima. Las medias de los valores de δ13C y δ18O de los individuos fueron comparadas entre ellos con el objeto de observar variaciones en la dieta a nivel población usando para ello un análisis de varianza (ANOVA) y la prueba de Tukey-Kramer para encontrar grupos estadísticamente distintos entre sí (Hammer y Harper, 2006).
Asimismo, en el caso de los valores de δ13C, se procedió a determinar el porcentaje de plantas C4 consumido (%C4), usando la ecuación propuesta por Koch et al. (2004):
(100)δ13Cmuestra = (100-x)δ13C100%C3esmalte + (X)δ13C100%C4esmalte
donde el valor de δ13C100%C3esmalte es de -12.5 ‰ y el de δ13C100%C4esmalte de 2.5‰, que corresponden a los estimados para el Pleistoceno Tardío (Koch, 2007).
El nivel de significancia fue establecido como p < 0.05 y los análisis fueron calculados usando el programa NCSS 2004 and PASS 2002 (Hintze, 2004), así como Excel 2003 (Microsoft Corporation, 2003).
4. Resultados
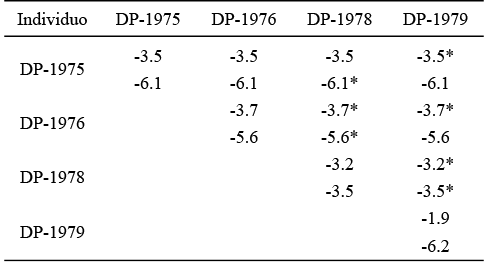
En las Tablas 1 y 2 se pueden observar los resultados obtenidos en los análisis isotópicos de δ13C y δ18O de cada uno de los individuos muestreados. En el caso del individuo DP-1975 los valores promedio de δ13C y δ18O son de -3.5 ‰ y -5.7 ‰, respectivamente. La Figura 3 muestra que los valores de δ13C se mantienen constantes al pasar de -3.5 ‰ a -3.6 ‰; sucede lo mismo con la δ18O que va de -5.9 ‰ a -6.3 ‰. Para el individuo DP-1976, la δ13C promedio fue de -2.7 ‰ y para la δ18O fue de -5.57 ‰, observándose que en las tres primeras los valores de δ13C se mantienen relativamente constantes, de -3.5 ‰ a -3.5 ‰ y posteriormente a -3.9 ‰, y volviéndose a mantener sin cambios abruptos para concluir en -3.9 ‰. Los valores isotópicos de oxígeno muestran que de la primera placa hacia la quinta placa se mantienen relativamente constantes y después de ésta, los valores de δ18O comienzan a aumentar al pasar de -5.9 ‰ a -4.8 ‰ (Figura 4).
La primera placa del individuo DP-1978 mostró una media de -3.2 ‰ para la δ13C y de -3.4 ‰ para la δ18O; en la Figura 5 se puede ver que el valor de δ13C sufre un descenso al pasar de -2.6 ‰, en la primera muestra, a -3.1 ‰, en la segunda, y posteriormente los valores isotópicos de carbono se mantienen constantes hasta la cuarta muestra, donde pasan de -3.1 ‰ a -3.5 ‰ en la quinta muestra, manteniéndose este valor en la última muestra. En el caso de los valores de la δ18O se observa un descenso abrupto del valor de la primera muestra a la segunda, de -1.9 ‰ a -3.7 ‰ y posteriormente pasa a -3.5 ‰, volviendo el valor isotópico de oxígeno a descender a -4.1 ‰ y manteniéndose relativamente constante en la quinta muestra y después descender a -3.8 ‰ en la última.
En la segunda placa del mismo individuo el valor promedio de δ13C es de -3.2 ‰ y la de δ18O fue de -3.5 ‰; sin embargo, a diferencia de la anterior placa, los valores isotópicos de carbono disminuyen constantemente, de -2.5 ‰ en la primera muestra a -3.8 ‰ en la última (Figura 6). Esta misma tendencia se observa con los valores de δ18O, pasando de -2.8 ‰ de la primera muestra a -4.9 ‰ en la sexta muestra y descendiendo a -4.1 ‰ en la última. En cambio, el individuo DP-1979 exhibe valores promedios de δ13C de -1.9 ‰ y de -6.2 ‰ para la δ18O. La Figura 7 indica que el valor de δ13C comienza a aumentar constantemente al pasar de -2.1 ‰, en la primera muestra, a -1.6 ‰, en la octava; una tendencia similar se observa en los valores de δ18O, en la primera muestra éste es de -6.8 ‰ y en la última fue de -5.0 ‰.
El ANOVA realizado entre los valores de δ13C y δ18O de los ejemplares, muestra que existen diferencias significativas para los valores de δ13C (p < 0.000001), al igual que los de δ18O (p < 0.000001). La prueba de Tukey-Kramer indica que el valor de δ13C del individuo DP-1975 es estadísticamente diferente al del DP-1979 y el valor de este isótopo en el individuo DP-1976 es diferente a los de DP-1978 y DP-1979. Asimismo el valor isotópico de oxígeno del DP-1975 es distinto al que presenta el DP-1978, mientras que el valor de δ18O del DP-1976 es diferente con respecto al mostrado por el individuo DP-1978. De manera similar el valor de δ18O de este último individuo es diferente al que se encuentra en el individuo DP-1979 (Tabla 3).

Figura 3. Valores isotópicos de carbono (rombos) y de oxígeno (cuadrados) del individuo DP-1975. Los valores de VPDB están expresados en δ y ‰.
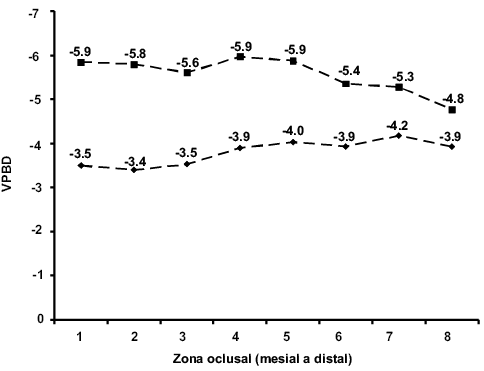
Figura 4. Valores isotópicos de carbono (rombos) y de oxígeno (cuadrados) del individuo DP-1976. Los valores de VPDB están expresados en δ y ‰.
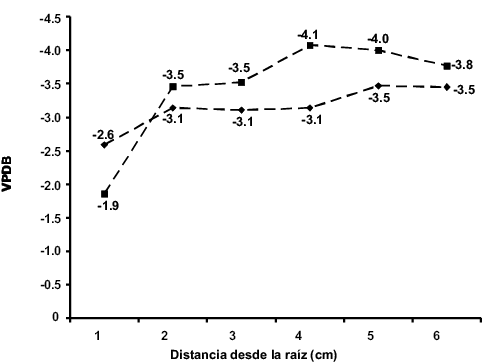
Figura 5. Valores isotópicos de carbono (rombos) y de oxígeno (cuadrados) de la primera placa del individuo DP-1978. Los valores de VPDB están expresados en δ y ‰.
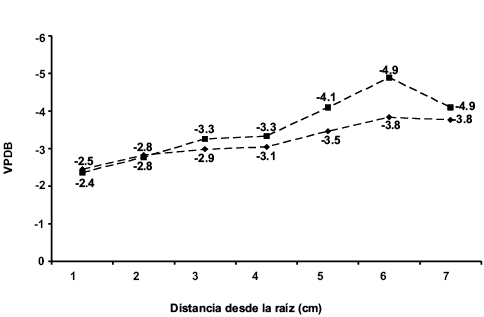
Figura 6. Valores isotópicos de carbono (rombos) y de oxígeno (cuadrados) de la segunda placa del individuo DP-1978. Los valores de VPDB están expresados en δ y ‰.
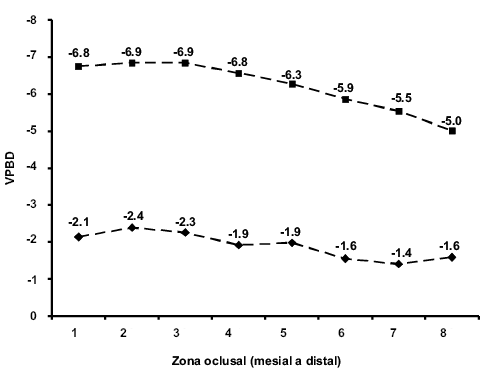
Figura 7. Valores isotópicos de carbono (rombos) y de oxígeno (cuadrados) del individuo DP-1979. Los valores deVPDB están expresados en δ y ‰.
Tabla 1. Datos obtenidos en los análisis del laboratorio de los Mammuthus columbi de Laguna de las Cruces. *Datos de la segunda placa del individuo DP-1978.
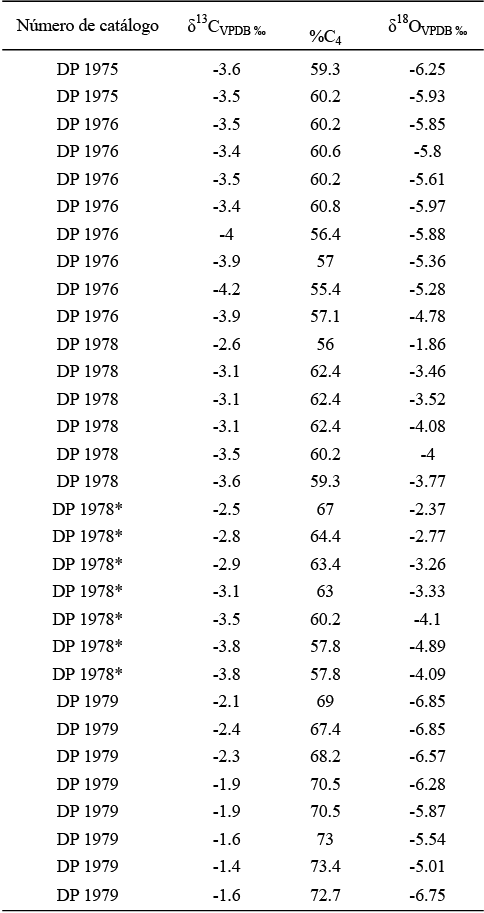
Tabla 2. Estadísticos descriptivos. P: número de placas muestreadas; +: muestras obtenidas de la zona oclusal por cada placa; ^: muestras obtenidas de la unión raíz-esmalte hacia la zona oclusal de una o dos placas; *: valores de ambas placas del DP 1978; n: número de muestras obtenidas. Los valores de carbono y oxígeno están expresados en VPDB‰.
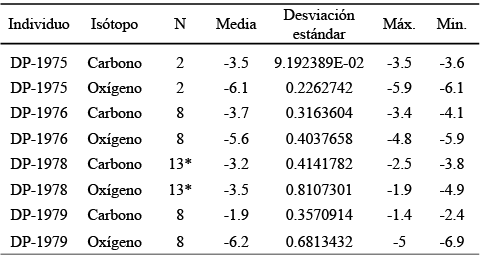
Tabla 3. Resultado de la prueba de Tukey para carbono y oxígeno (segunda línea). * Grupos que son diferentes.
5. Discusión
La dieta promedio del individuo DP-1975 muestra una alimentación mixta C3/C4 (%C4: 59.8), la cual se mantuvo constante durante la formación de ambas placas y que se realiza alrededor de 43 años (%C4: 59 a 60.2) al igual que la temperatura (Dirks et al., 2012). En cambio, en el individuo DP-1976, cuya dieta también fue del tipo mixta C3/C4, se observa una ligera disminución en el consumo de plantas C4 y un aumento en la ingesta de plantas C3, sin llegar a ser ésta una parte importante de su dieta, al pasar de un %C4 de 60 en la primera muestra a un %C4 de 57.1 en la última; esto se relaciona con pequeñas disminuciones en la temperatura ambiente.
Una tendencia similar se puede observar en las dos placas muestreadas del individuo DP-1978, cuyo promedio de consumo de plantas C4 fue de 62 %; en la primera placa el consumo de plantas C4 fue de 56 %, aumentando a 62.4 %, manteniéndose ésta constante hasta la cuarta muestra, disminuyendo en la quinta a 60.2% y finalmente acabando en 59.3 % En el caso de la segunda placa la dieta comienza con un %C4 de 67 y finaliza con un %C4 de 57.2; los valores de oxígeno muestran un decremento en la temperatura que pudiera ocasionar una disminución de las plantas C4 y el aumento en el consumo de las plantas C3 en la dieta de este individuo, conforme ambas placas se formaban.
En contraste, para el DP-1979, el tipo de dieta fue completamente pacedora (%C4: 70.6); sin embargo, al inicio de la formación de la placa el individuo tenía una dieta mixta C3/C4 (%C4: 69), pero el consumo de las plantas C3 comienza a disminuir y el de las C4 a aumentar hasta que estas llegan a ser su principal fuente de alimento (%C4: 72.7).
Dado que la temperatura es uno de los factores que controla la distribución y la abundancia de las plantas C3 y C4 (Medrano y Flexas, 2000), se puede observar en los casos anteriores que el aumento o el descenso en el consumo de un tipo de planta va asociado a la variaciones en la temperatura, ya que a temperaturas mayores de 25 ºC las plantas C4 abundan y a temperaturas menores a 25 ºC son las C3 las que predominan (Feranec y MacFadden, 2000).
Haynes y Klimowicz (2003) han indicado que Mammuthus columbi tenía una conducta parecida a la de los elefantes actuales, los cuales poseen una conducta alimenticia intraespecífica distinta debido tanto al sexo como a la edad. Sukumar et al. (1987), Sukumar y Ramesh (1992) y Greyling (2004) han encontrado que en los elefantes africanos y asiáticos los adultos tienden a ramonear más que los jóvenes, los cuales prefieren consumir pastos principalmente. Usando la metodología de Laws (1966) se obtiene que el individuo DP-1978 tenía una edad de 20 ± 1 años y el DP-1979 de 18 ± 1 años y los DP-1975 y DP-1976 edades de 43 ± 2 años y 47 ± 2 años, respectivamente y, dado que los valores isotópicos de carbono de los individuos juveniles son semejantes entre ellos pero diferentes los de los adultos, es posible que este tipo de conducta alimentaria se refleje en los valores de δ13C de los mamuts.
Otra posible explicación es que no todos los individuos eran nativos de la zona; en el caso de los elefantes, Haynes (1991) menciona que son capaces de desplazarse hasta 100 km en busca de comida y agua, por lo que constantemente se trasladan por diferentes zonas y manadas distintas pueden usar las mismas fuentes de agua. Esto posiblemente explique las variaciones en los valores de δ18O observadas en las placas de cada uno de los individuos, quienes pudieron estar en distintos sitios y solo acudían a beber al cuerpo de agua que estaba presente en el sitio durante el Pleistoceno Tardío. Datos preliminares usando las relaciones de 87Sr/86Sr muestran las mismas diferencias halladas en los valores de δ13C y δ18O de estos individuos, lo cual podría apoyar lo anterior (Solís-Pichardo et al., 2011); sin embargo, la falta de valores isotópicos provenientes de los suelos y las plantas del área hace difícil señalar qué individuos eran nativos de la zona y cuáles provenían de otras.
6. Conclusión
Los análisis isotópicos realizados en los mamuts de Laguna de las Cruces, indican que existen variaciones en el consumo de las plantas C3 y C4 de los individuos muestreados. Así, en el caso del individuo DP-1975 ésta se mantuvo constante, mientras que en los individuos DP-1976 y DP-1978, hubo un ligero aumento en el consumo de plantas C3 debido a que la temperatura disminuyó ligeramente. En cambio en el individuo DP-1979 su ingesta de plantas C3 comenzó a disminuir hasta que éstas desaparecieron de su dieta y sólo se alimentó de plantas C4, por un aumento de la temperatura en la zona. Asimismo, los análisis isotópicos indican que existe una diferencia entre el tipo de alimentación que puede ser explicada por la conducta alimentaria que presentan los dos individuos juveniles, la cual es diferente a la que exhiben los individuos adultos, pero no se puede descartar que algunos individuos sean residentes de la zona y que otros provengan de otras sitios.
Agradecimientos
Al Consejo Nacional de Arqueología por permitir tomar las muestras esmalte de los mamuts de la Colección de Paleontología del Instituto Nacional de Antropología e Historia. A CONACYT por su apoyo para completar parte del estudio, a través del proyecto “Partición de Recursos de los Mega y Mesoherbívoros del Pleistoceno Tardío de El Cedral y Laguna de las Cruces, San Luis Potosí, México” (Convenio CB-2009-01-132920). Al CONACyT y el Instituto Politécnico Nacional por las becas de estudios de maestría 223602 SIP20050200 y SIP20060322, respectivamente. Al Laboratorio Universitario de Geoquímica Isotópica (LUGIS) del Instituto de Geología de la UNAM y a R. Puente M. por analizar las muestras. A los Drs. Maria Teresa Alberdi, Federico Agnolin y José Luis Padro por sus comentarios que enriquecieron el presente manuscrito.
Referencias
Álvarez, T., 1982, Restos óseos animales localizados en las excavaciones, en Mirambell, L. (ed.), Laguna de las Cruces, Salinas, S. L. P. Un sitio paleontológico del Pleistoceno final: Colección Científica 128, Instituto Nacional de Antropología e Historia, México, 62-75.
Andrade, J.L., de la Barrera, E., Reyes-García, C., Ricalde, M.F., Vargas-Soto, G., Cervera. J.C., 2007, El metabolismo ácido de las crasuláceas: Boletín de la Sociedad Botánica de México, 81, 37-50.
Andrews, P., 1996, Palaeocology and hominoid palaeoenvironments: Biological Reviews, 71, 257-300.
Arroyo-Cabrales, J., Polaco, O.J., Laurito, C., Johnson, E., Alberdi, M.T., Valerio, A.L, 2007, The proboscideans (Mammalia) from Mesoamerica: Quaternary International, 169, 17-23.
Bell, C.J., Lundelius Jr., E.L., Barnosky, A.D., Graham, R.W., Lindsay, E.H., Ruez Jr., D.R., Semken Jr., H.A., Webb, S.D., Zakrzewski, R.J., 2004, The Blancan, Irvingtonian, and Rancholabrean mammal ages, en Woodburne, M.O. (ed.), Late Cretaceous and Cenozoic mammals of North America: New York,Columbia University Press, USA, 232-314.
Bravo-Cuevas. V.M., Jiménez-Hidalgo, E., Priego-Vargas, J, 2011, Taxonomía y hábitos alimentarios de Equus conversidens (Perissodactyla, Equidae) del Pleistoceno Tardío (Rancholabreano) de Hidalgo, centro de México: Revista Mexicana de Ciencias Geológicas, 28, 65-82.
Cerling, T.E., Harris, J.M., MacFadden, B.J., Leakey, M.G., Quade, J., Eisenmann, V., Ehleringer, J.R., 1997, Global vegetation change through the Miocene/Pliocene boundary: Nature, 389,153-158.
Cerling, T.E., Harris, J.M., 1999, Carbon isotope fractionation between diet and bioapatite in ungulate mammals and implications for ecological and paleoecological studies: Oecologia, 120, 347-363.
Coplen, T.B., 1988, Normalization of oxygen and hydrogen isotope data: Chemical Geology, 72, 293-297.
Coplen, T., Brand, W.A., Gehre, M., Gröning, M., Meijer Harro, A.J., Toman, B., Erkouteren, R.M., 2006, New Guidelines for δ13C Measurements: Analytical Chemistry, 78, 2439-2441.
Dawson, T.E., Mambelli, S., Plamboeck, A.H., Templer, P.H., Tu, K.P., 2002, Stable isotopes in plant ecology: Annual Review Ecology and Systematics, 33, 507-559.
DeNiro, M.J., Epstein, S., 1978, Influence of diet on the distribution of carbon isotopes in animals: Geochimica et Cosmochimica Acta, 42, 495-506.
Dirks, W., Bromage, T.G., Agenbroad, L.D., 2012, The duration and rate of molar plate formation in Palaeoloxodon cypriotes and Mammuthus columbi from dental histology: Quaternary International, 255, 79-85.
Ehleringer , J.R., Cerling, T.E., Helliker, B.R., 1997, C4 photosynthesis, atmospheric CO2, and climate: Oecologia, 112, 285-299.
Ericson, J.E., Sullivan, C.H., Boaz, N.T., 1981, Diets of Pliocene mammals from Omo, Ethiopia, deduced from carbon isotopic ratios in tooth apatite: Palaeogeography, Palaeoclimatology, Palaeoecology, 36, 69-73.
Feranec, R.S, MacFadden, B., 2000, Evolution of the grazing niche in Pleistocene mammals from Florida: evidence from stable isotopes: Palaeogeography, Palaeoclimatology, Palaeoecology, 162, 155-169.
Flores, A.D., 1982, Análisis físicos y químicos de la Laguna de las Cruces, en Mirambell, L. (ed.), Laguna de las Cruces, Salinas, S. L. P. Un sitio paleontológico del Pleistoceno final: Colección Científica 128, Instituto Nacional de Antropología e Historia, México, 35-58.
Fricke, H.C., O’Niel J.R., 1996, Inter- and intra-tooth variation in the oxygen isotope composition of mammalian tooth enamel phosphate: implications for palaeoclimatological and palaeobiological research: Palaeogeography, Palaeoclimatology, Palaeoecology, 126, 91-99.
Greyling, M.D., 2004, Sex and age related distinctions in the feeding ecology of the African elephant Loxodonta africana: Witwatersrand, Sudafrica, Universidad de Witwatersrand, tesis doctoral, 189 p.
Hammer, Ø., Harper, D., 2006, Paleontological Data Analysis: Boston, Blackwell Publishing, 351 p.
Haynes, G., 1991, Mammoths, Mastodonts and Elephants. Biology, Behavior and the Fossil Record: Nueva York, Cambridge University Press, 413 p.
Haynes, G., Klimowicz, J., 2003, Mammoth (Mammuthus spp.) and American mastodont (Mammut americanum) bonesites: what do the differences mean: DEINSEA, 9, 185-204.
Hintze, J., 2004, NCSS and PASS Number Cruncher Statically System (CD-ROM): Kaysville, UTHA, WWW. NCSS.COM., 1 CD-ROM.
Hofmann, R.R., Stewart, D.R.M., 1972, Grazer or browser: a classification based on stomach-structure and feeding habits of East African ruminants: Mammalia, 36, 226-240.
Keeley, J.E., Rundel, P.W., 2003, Evolution of CAM and C4 carbon-concentrating mechanisms: International Journal of Plant Science, 164 (suplemento 3), S55-S77.
Kingston, J.D., Harrison, T., 2007, Isotopic dietary reconstructions of Pliocene herbivores at Laetoli: Implications for early hominin paleoecology: Palaeogeography, Palaeoclimatology, Palaeoecology, 243, 272-306.
Koch, P.L., 1998, Isotopic reconstruction of past continental environments: Annual Review of Earth and Planetary Sciences, 26, 573-613.
Koch, P.L., 2007, Isotopic study of the biology of modern and fossil vertebrates, en Michener, R., Lajtha, K. (eds.), Stable Isotopes in Ecology and Environmental Science: Boston, Blackwell Scientific Publications, 99-154.
Koch, P.L., Behrensmeyer, A.K., Fogel, M.L., 1991, The isotopic ecology of plant and animals in Ambroseli National Park, Kenya, en Prewitt, C.T. (ed.), Annual Report of the Director of the Geophysical Laboratory, Carnegie Institution of Washington: Washington, D.C., Carnegie Institution of Washington, 163-171.
Koch, P.L., Fogel, M.L., Tuross, N., 1994, Tracing the diets of fossil animals using stable isotopes, en Lajtha, K., Michener, R.H. (eds.), Stable Isotopes in Ecology and Environmental Science: Boston, Blackwell Scientific Publications, 63-92.
Koch, P.L., Tuross, N., Fogel, M.L., 1997, The effects of sample treatment and diagenesis on the isotopic integrity of carbonate in biogenic hydroxylapatite: Journal of Archaeological Science, 24, 417-429.
Koch, P.L., Diffenbaugh, N.S., Hoppe, K.A., 2004, The effects of late Quaternary climate and PCO2 change on C4 plant abundance in the south-central United States: Palaeogeography, Palaeoclimatology, Palaeoecology, 207, 331-357.
Laws, R.M., 1966, Age criteria for the African elephant Loxodonta africana: East African Wildlife Journal, 4, 1-37.
Longinelli, A., Nuti, S., 1973, Revised phosphate-water isotopic temperature scale: Earth and Planetary Science Letters, 19, 373-376.
MacFadden, B.J., Cerling, T.E., 1996, Mammalian herbivore communities, ancient feeding ecology, and carbon isotopes: a 10 million-year sequence from the Neogene of Florida: Journal of Vertebrate Paleontology, 16, 103-115.
MacFadden, B.J., Wang, Y., Cerling, T. E., Anaya, F., 1994, South American fossil mammals and carbon isotopes: a 25 million-year sequence from the Bolivian Andes: Palaeogeography, Palaeoclimatology, Palaeoecology, 107, 257-268.
MacFadden, B.J., Cerling, T., Harris, J.M., Prado, J., 1999, Ancient latitudinal gradients of C3/C4 grasses interpreted from stable isotopes of New World Pleistocene horse (Equus) teeth: Global Ecology and Biogeography, 8, 137-149.
Medrano, H., Flexas, J., 2000, Fotorrespiración y mecanismos de concentración del dióxido de carbono, en Azcón-Bieto, J.,Talón. M. (eds.), Fundamentos de fisiología vegetal: Madrid, McGraw-Hill Interamericana, 187-201.
Metcalfe, J.Z., Longstaffe, F. J., 2012, Mammoth tooth enamel growth rates inferred from stable isotopes análisis and histology: Quaternary Research, 77, 424-432.
Microsoft Corporation, 1985- 2003, Excel 2003 (CD-ROM): Redmon, Washington, Microsoft Corporation, 1 CD-ROM.
Mirambell, L., 1982, Las excavaciones, en Mirambell, L., (ed.), Laguna de las Cruces, Salinas, S. L. P. Un sitio paleontológico del Pleistoceno final: Colección Científica 128, Instituto Nacional de Antropología e Historia, México, 12-18.
Montellano-Ballesteros, M, Jiménez-Hidalgo, E., 2006, Mexican fossil mammals, Who, where and when? en Vega, J.F., Nyborg, T.G., Perrilliat, M.C., Montellano-Ballesteros, M., Cevallos-Ferriz, S.R.S., Quiroz-Barroso, S.A. (eds.), Studies on Mexican Paleontology: Nueva York, Springer, 249-273.
O’Leary, M.H., 1981, Carbon isotope fractionation in plants: Phytochemistry, 2, 553-567.
Pérez-Crespo, V.A., Sánchez-Chillón, B., Arroyo-Cabrales, J., Alberdi, M.T., Polaco, O.J., Santos-Moreno, A., Benammi, M., Morales-Fuentes, P., Cienfuegos-Alvarado, E., 2009, La dieta y el hábitat del mamut y los caballos del Pleistoceno tardío de El Cedral con base en isótopos estables (δ13C, δ18O): Revista Mexicana de Ciencias Geológicas, 26, 347-355.
Pérez-Crespo, V.A., Arroyo-Cabrales, J., Benammi, M., Santos-Moreno, A., Morett-A, L., Morales-Puente, P., Cienfuegos-Alvarado, E., 2010, Carbon isotopic values of tooth enamel of Mammuthus columbi from Tocuila, State of México, México: Current Research in the Pleistocene, 27, 188-190.
Pérez-Crespo, V.A., Arroyo-Cabrales, J., Alva-Valdivia, L.M., Morales-Puente, P., Cienfuegos-Alvarado, E., 2012, Datos isotópicos (δ13C, δ18O) de la fauna pleistocenica de la Laguna de las Cruces, San Luis Potosí, México: Revista Mexicana de Ciencias Geológicas, 29, 299-307.
Révész, K.M., Landwehr, J.M., 2002, δ13C and δ18O isotopic composition of CaCO3 measured by continuous flow isotope ratio mass spectrometry: statistical evaluation and verification by application to Devils Hole core DH-11 calcite: Rapid Communications in Mass Spectrometry, 16, 2102-2114.
Reyes, C.M., 1982, Consideraciones sobre la geología del Cuaternario del área, en Mirambell, L. (ed.), Laguna de las Cruces, Salinas, S. L. P. Un sitio paleontológico del Pleistoceno final: Colección Científica 128, Instituto Nacional de Antropología e Historia, México, 22-31.
Sánchez, B., 1997, Estudio de las variaciones climáticas durante el final del Neógeno a partir del análisis de δ18O (PO43-) en fósiles de mamíferos, en Calvo, J.P., Morales, J. (eds.), Avances en el conocimiento del Terciario Ibérico: Madrid, Universidad de Madrid, 197-200.
Sánchez, B., 2005, Reconstrucción del ambiente de mamíferos extintos a partir del análisis isotópico de los restos esqueléticos, en Alcorno, P., Redondo, R., Toledo, J. (eds.), Nuevas técnicas aplicadas al estudio de los sistemas ambientales: los isótopos estables: Madrid, Universidad Autónoma de Madrid, 49-64.
Solís-Pichardo, G., Pérez-Crespo, V. A., Schaaf, P., Arroyo-Cabrales, J., 2011, Sr isotopes and migration of praire mammoths (Mammuthus columbi) from Laguna de las Cruces, San Luis Potosí, México (resumen), en Joint Assembly of American Geophysical Union, San Francisco California, USA, 5-9.
Smith, B.N., Epstein, S., 1971, Two categories of 13C/12C ratios for higher plants: Plant Physiology, 47, 380-384.
Stuart-Williams, H.L., Schwarcz, H.P., 1997, Oxygen isotopic determination of climatic variation using phosphate from beaver bone, tooth enamel, and dentine: Geochemica et Cosmochimica Acta, 61, 2539-2550.
Sukumar, R., Ramesh, R., 1992, Stable carbon isotope ratios in Asian elephant collagen: implications for dietary studies: Oecologia, 192, 536-539.
Sukumar, R., Bhattacharya, S.K., Krishnamurthy, R.V., 1987,Carbon isotopic evidence for different feeding patterns in an Asian elephant population: Current Science, 56, 11-14.
Van der Merwe, N. J., Medina, E., 1989, Photosynthesis and 13C/12C ratios in Amazonian rain forest: Geochimica et Cosmochimica Acta, 53,1091-1094.
Werner, R.A., Brand, W.A., 2001, Referencing strategies and techniques in stable isotope ratio analysis: Rapid Communications in Mass Spectrometry, 15, 501-519
Manuscrito recibido: Diciembre 18, 2012.
Manuscrito corregido recibido: Marzo 12, 2013.
Manuscrito aceptado: Abril 7, 2013.