
|
BOLETÍN DE LA SOCIEDAD GEOLÓGICA MEXICANA Vol 65, Núm. 3, 2013, P. 533-551 http://dx.doi.org/10.18268/BSGM2013v65n3a8 |
 |
Bivalvos guadalupianos (Pérmico medio) de la Formación Olinalá, Guerrero, México
Miguel Guerrero-Sánchez1, Sara A. Quiroz-Barroso2,*
1Facultad de Ciencias, Universidad Nacional Autónoma de México, Ciudad Universitaria, 04510 México D.F., México.
2Museo de Paleontología, Departamento de Biología Evolutiva, Facultad de Ciencias, Universidad Nacional Autónoma de México, Ciudad Universitaria, 04510 México D.F., México.
* This email address is being protected from spambots. You need JavaScript enabled to view it.
Resumen
El material estudiado proviene de rocas que afloran al noreste del poblado Olinalá, en el estado de Guerrero. Las facies marinas de la Formación Olinalá están incluidas en lutita gris claro a oscuro con fósiles de braquiópodos, gasterópodos, bivalvos, cefalópodos, tallos de crinoideos, madera y otros restos de plantas, localmente alojados en concreciones. Entre la diversidad de los bivalvos se encuentran las especies infaunales Nuculavus levatiformis, Palaeoneilo mcchesneyana, Edmondia phosphatica y ?Cardiomorpha sp.; las especies epifaunales que vivían adheridas a diferentes sustratos Posidoniella sp., Septimyalina burmai y Leptodesma (Leptodesma) sp.; así como la especie semi-infaunal Bakevellia sp. Los hábitos de vida de los bivalvos y las características de su preservación indican que fueron poco transportados antes de su depósito final, lo cual ocurrió en una plataforma de aguas poco profundas. Las especies de bivalvos estudiadas concuerdan con la edad pérmica de esta formación, establecida previamente por otros autores con base en el estudio de amonites y foraminíferos, y apoyan la idea de su relación con faunas contemporáneas de América del Norte.
Palabras clave: Formación Olinalá, bivalvos, Pérmico medio, México.
Abstract
The study material comes from rocks that outcrop northeast of the town Olinalá, in Guerrero state. The marine facies of Olinalá Formation are included in light to dark gray shale with fossils of brachiopods, gastropods, bivalves, cephalopods, crinoids stems, wood and other plant remains, locally included in concretions. The diversity of bivalves is represented by the infaunal species Nuculavus levatiformis, Palaeoneilo mcchesneyana, Edmondia phosphatica, and ?Cardiomorpha sp.; the epifaunal species, adhered to different substrates Posidoniella sp., Septimyalina burmai, and Leptodesma (Leptodesma) sp.; and the semi-infaunal species Bakevellia sp. The lifestyle and preservation characteristics of bivalves indicate that they were shortly transported before its final deposition, which occurred on a platform, in shallow water. The studied bivalve species are consistent with the permian age of this formation, considered previously by other authors based on the study of ammonites and foraminifera, and support the idea of their relationship with contemporary faunas of North America.
Palabras clave: Olinalá Formation, bivalves, middle Permian, Mexico.
1. Introducción
Las rocas que afloran en la parte noreste del poblado Olinalá, en el estado de Guerrero, constituyen una sucesión de más de 800 m de espesor que ha sido denominada como Formación Los Arcos (Corona-Esquivel, 1981 [1983]) y Formación Olinalá (Flores de Dios y Buitrón, 1982), siendo este último nombre el reconocido como válido en el Léxico Estratigráfico del Servicio Geológico Mexicano (SGM, 2006).
La Formación Olinalá está constituida por una diversidad de facies terrígenas (marinas y continentales) y carbonatadas (marinas), cuyos ambientes de depósito varían de fluviales a margen de rampa, costeros y plataforma externa (Juárez-Arriaga, 2006). De ellas, las facies marinas de lutita son las más abundantes tanto en la base como en la cima de la formación y consisten en lutita gris claro a oscuro finamente laminada, con alto contenido de biotita en la base y abundantes concreciones, generalmente paralelas a la estratificación (Juárez-Arriaga, 2006). En cuanto a su fauna se ha mencionado la presencia de calcimicrobios (Tubiphytes sp.), foraminíferos, briozoarios, braquiópodos, bivalvos, gasterópodos, amonoideos, artrópodos (ostrácodos y trilobites), crinoideos, así como restos vegetales (Corona-Esquivel, 1981 [1983], 1985; Flores de Dios y Buitrón, 1982; Flores de Dios, 1986; González-Arreola et al., 1994; Silva-Pineda et al., 1998; Flores de Dios et al., 2000; Garibay-Romero, 2000; Esquivel-Macías, 2000; Esquivel-Macías et al., 2004; Vachard et al., 2004; Juárez-Arriaga, 2006).
La presencia de bivalvos en la facies de lutita marina se menciona e ilustra en los estudios de González-Arreola et al. (1994) y Juárez-Arriaga (2006). En el presente trabajo se describen las especies de bivalvos de la lutita media de esta formación y se propone una interpretación acerca de su presencia en rocas del Paleozoico superior de México, analizándose sus hábitos de vida, así como su distribución geográfica y estratigráfica.
2. Ubicación del área de estudio
El material estudiado se recolectó en rocas que afloran al noreste de Olinalá, en el estado de Guerrero (Figura 1).
Localidad 1. Sobre el camino de terracería que parte desde Olinalá hacia el poblado El Progreso, con coordenadas 17° 46.864’ latitud N y 98° 43.761’ longitud W. Lutita de color gris oscuro que intemperiza en forma de láminas delgadas, con manchas color ocre debidas a la oxidación y con contenido fósil. Contiene concreciones que varían en tamaño de 1.5 a 20 cm, en muchas de las cuales hay contenido fósil; algunas de las concreciones son alargadas y se encuentran sobre el plano de estratificación. Se presentan moldes de ejemplares grandes del bivalvo Posidoniella De Koninck, 1885, así como moldes de otros bivalvos (Septimyalina Newell, 1942; Bakevellia King, 1848 y Leptodesma Hall, 1883), gasterópodos, amonoideos y un fragmento de un tallo de crinoideo. Se observan impresiones de restos vegetales.
Localidad 2. Sobre el camino de terracería que parte desde Olinalá hacia el poblado El Progreso, con coordenadas 17° 47.160’ latitud N y 98° 43.861’ longitud W. Lutita negra con estratificación paralela. El echado de las capas es de casi 90°. Están presentes moldes de braquiópodos, bivalvos (Nuculavus Chernyshev, 1947; Palaeoneilo Hall y Withfield, 1869; Posidoniella, Septimyalina, Edmondia De Koninck, 1841 y ?Cardiomorpha De Koninck, 1844), gasterópodos, nautiloideos y placas de crinoideos.
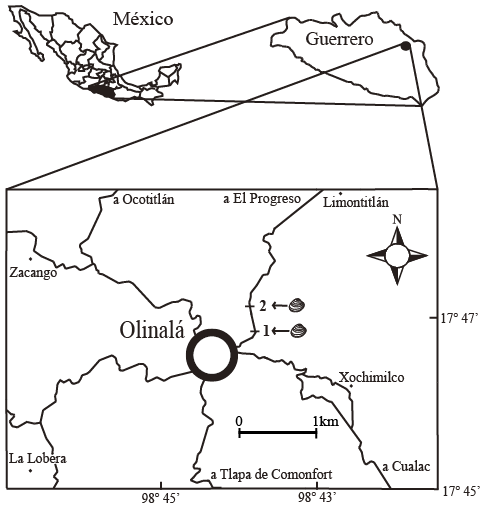
Figura 1. Área de estudio. Las barras negras con números representan la ubicación de las localidades de recolecta
3. Estratigrafía
Las rocas de la Formación Olinalá yacen discordantemente sobre rocas metamórficas del Complejo Acatlán de edad Ordovícico Tardío-Devónico Tardío (Ortega-Gutiérrez et al., 1999) y están cubiertas discordantemente por la Ignimbrita Las Lluvias de edad Triásico?-Jurásico Medio (Corona-Esquivel, 1981 [1983]; García-Díaz et al., 2000). A su vez, esta unidad está cubierta por el Conglomerado Cualac y el Grupo Tecocoyunca (Erben, 1956) del Jurásico Medio.
Corona-Esquivel (1981 [1983]), reconoció siete unidades en la Formación Olinalá. La unidad uno es la base de la formación y es un conglomerado poligenético conformado por guijarros sub-redondeados de esquisto de mica, cuarzo blanco, cuarcita, esquisto verde de clorita y algunos fragmentos de filita. La unidad dos consiste en limolita negra y lutita gris oscura a negra, con algunas capas de arenisca gris de grano fino, de 15 cm de espesor y abundantes concreciones limolíticas; en la base se presenta un horizonte con amonites y bivalvos dentro de las concreciones. La unidad tres se divide en dos partes: a) una inferior, que consiste en capas gruesas de conglomerado poligenético con guijarros de gneis sub-angulosos y redondeados, esquisto, pizarra negra y cuarcita gris claro; y b) una superior con arenisca de grano medio a grueso de color gris claro, en capas de 0.20 a 0.80 m de espesor, muy resistentes. La unidad cuatro presenta limolita a lutita de color gris oscuro a negro, con estratificación no definida y con ligera foliación. La unidad cinco consiste en caliza gris que intemperiza a gris claro y su espesor varía regionalmente; presenta abundantes crinoideos y gasterópodos. La unidad seis se define como una alternancia de arenisca de color café claro en capas medianas a delgadas y lutita de color gris oscuro con intercalaciones de horizontes con concreciones calcáreas. La unidad siete es la más joven y consiste en una arenisca de color negro a gris oscuro con estratificación delgada.
De acuerdo con Juárez-Arriaga (2006) las facies continentales de la Formación Olinalá están compuestas por arenisca conglomerática, conglomerado polimíctico y cuarzo-arenita que se interpretan como ambientes de depósito fluvial o costero. El mismo autor subdivide las facies marinas en carbonatadas, que incluyen biohermas de algas rojas coralináceas, packstone arcilloso de braquiópodos y wackestone/packstone de fusulínidos, packstone/grainstone de crinoideos-briozoarios-braquiópodos, entre otros; y terrígenas, compuestas por lutita de color negruzco, laminación fina, con pirita, estructuras cono-en-cono, y fósiles tales como amonoideos, nautiloideos, gasterópodos, braquiópodos, conuláridos, fragmentos desarticulados de crinoideos, restos vegetales, además de bivalvos. Por sus relaciones estratigráficas, sugieren un depósito en condiciones de plataforma externa, sin influencia de oleaje. Los ciclos en el depósito han sido considerados por el mismo autor, como el registro de tres eventos transgresivos-regresivos que ocurrieron en el área durante el Pérmico medio.
El material de bivalvos examinado se encuentra en estratos que corresponden a parte de la unidad 4 propuesta por Corona-Esquivel (1981 [1983]), y a la lutita de la secuencia SD-2 de Juárez-Arriaga (2006) (Figura 2).

Figura 2. Columna estratigráfica simplificada de la Formación Olinalá. Se indica la parte de la columna que corresponde a la unidad 4 de Corona-Esquivel (1981) y González-Arreola et al. (1994) del cual provienen los bivalvos estudiados. Modificada de Juárez-Arriaga (2006).
4. Consideraciones estratigráficas
Flores de Dios y Buitrón (1982) asignaron inicialmente a toda la Formación Olinalá una edad Pensilvánico-Triásico. Posteriormente Corona-Esquivel (1981 [1983]), Vachard et al. (1993), González-Arreola et al. (1994), Esquivel-Macías (1996) y Silva-Pineda et al. (1998, 2000) restringieron la edad al Pérmico.
Vachard et al. (2004) reconocieron una posible edad Wordiano para las capas inferiores de la Formación Olinalá basados en la presencia del amonoideo Waagenoceras sp., mientras que asignaron una edad Capitaniano a la parte carbonatada de esta formación por la presencia de los fusulínidos Polydiexodina sp. y Codonofusiella extensa; sin embargo, el alcance conocido de Polydiexodina se ha extendido, en los últimos años, del Wordiano al Capitaniano. La especie Codonofusiella extensa sólo se conoce para el Capitaniano de Coahuila, por lo que al tenerse un registro tan limitado de su distribución geográfica, tampoco puede ser considerada en este momento como un fósil índice; mientras que el género Waagenoceras presenta una distribución temporal conocida que va del Roadiano al Capitaniano.
De acuerdo a las consideraciones anteriores y a los alcances estratigráficos de los bivalvos estudiados (Figura 3), el conjunto es característico del Pérmico medio y sugiere una edad de Roadiano, lo que ampliaría la extensión del alcance estratigráfico de la especie Septimyalina burmai. Si este fuera el escenario real, habría importantes implicaciones biogeográficas, ya que en la Formación Olinalá existen especies de braquiópodos que han sido consideradas como probables formas endémicas relictas (Sour-Tovar et al., 2009).
Por otra parte, González-Arreola et al. (1994) propusieron ampliar el alcance estratigráfico de Posidoniella, un género previamente considerado del Carbonífero, hasta el Pérmico medio y posteriormente, Quiroz-Barroso et al. (2012) mencionaron la presencia de este género en rocas del Cisuraliano (Kunguriano) de la Formación Tuzancoa, en el estado de Hidalgo; con base en los resultados obtenidos en el presente estudio, se corrobora la propuesta de ampliar el alcance estratigráfico de este género hasta el Pérmico medio, por lo menos hasta el Roadiano.
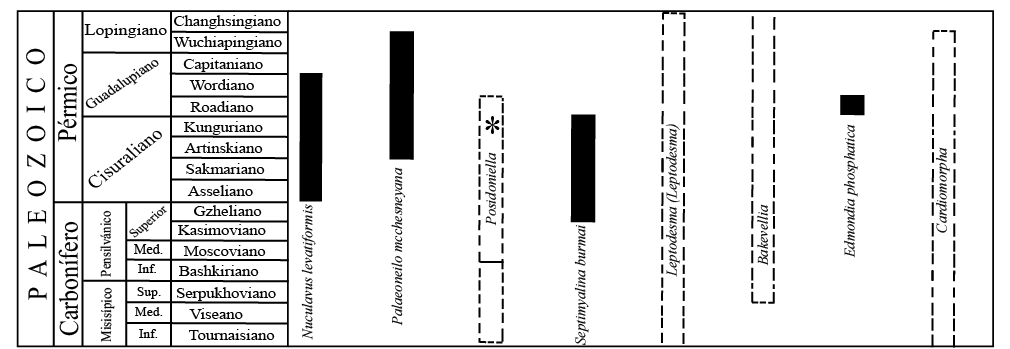
Figura 3. Alcances estratigráficos de los géneros y especies estudiados. Las líneas punteadas indican el alcance estratigráfico del género; las barras negras indican el alcance estratigráfico de las especies. En el caso de Posidoniella sp., la sección con asterisco (*) indica la propuesta de González-Arreola et al. (1994) de ampliar su alcance estratigráfico hasta el Pérmico medio.
5. Consideraciones tafonómicas y paleoecológicas
Los géneros de bivalvos estudiados representan tres modos de vida principales: Nuculavus, Palaeoneilo, Edmondia y Cardiomorpha incluyen especies que fueron organismos móviles facultativos y formaron parte de la infauna; Posidoniella, Septimyalina y Leptodesma incluyen especies que vivían adheridas a diferentes sustratos formando parte de la epifauna y Bakevellia es un género con especies semi-infaunales (Figura 4). Su registro consiste en moldes internos, moldes externos y vaciados en lutita obscura, cuya preservación fue favorecida, en muchos, casos al quedar incluidos en dos tipos de concreciones.
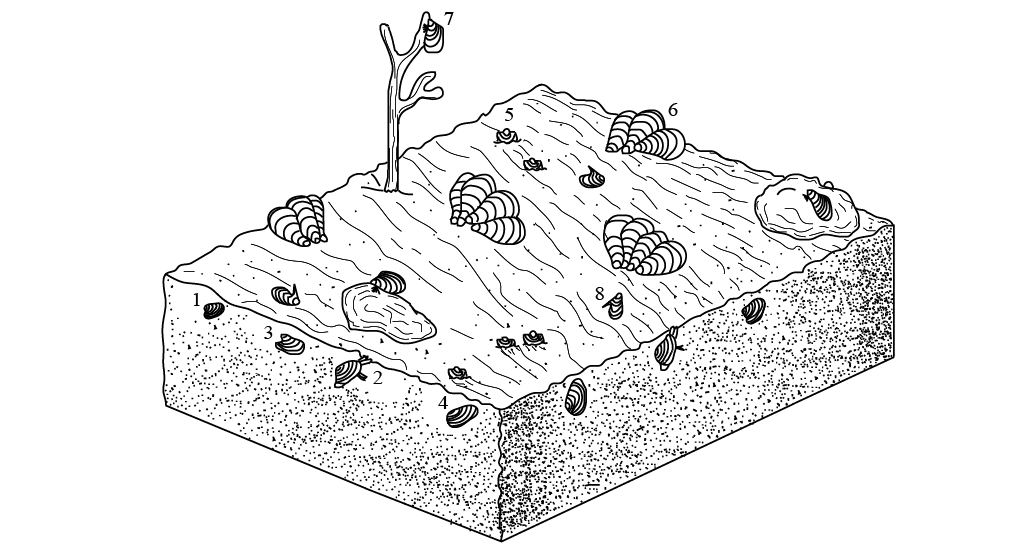
Figura 4. Hábitos de vida de los bivalvos estudiados. Infaunales, móviles facultativos: 1, Nuculavus levatiformis (detritívoro-suspensívoro); 2, Palaeoneilo mcchesneyana (detritívoro); 3, Edmondia phosphatica (suspensívoro); 4, Cardiomorpha sp. (suspensívoro). Semi-infaunal, estacionario: 5, Bakevellia sp. (suspensívoro). Epifaunales, estacionarios: 6, Posidoniella sp. (suspensívoro); 7, Septimyalina burmai (suspensívoro); 8, Leptodesma (Leptodesma) sp. (suspensívoro). No se trata de una reconstrucción ambiental.
Tipo 1. Las que tienen concentraciones de individuos de los géneros Nuculavus y Palaeoneilo, que fueron organismos infaunales de talla pequeña y cuyo registro en aguas marinas incluye facies de costa y de plataforma (Newell et al., 1953; Sterren, 2000; Anelli et al., 2002) (Figura 5). De acuerdo a la clasificación de Kidwell et al. (1986) las concentraciones observadas en este tipo de concreciones son politípicas por la presencia de varias especies de bivalvos y braquiópodos. Su estructura bioclástica consiste en conchas pequeñas, articuladas, dispuestas en forma azarosa y sin deformación por compresión.

Figura 5. Concreciones con conchas pequeñas de braquiópodos y de los bivalvos Nuculavus levatiformis (1) y Palaeoneilo mcchesneyana (1 y 2).
Tipo 2. Aquellas en las que hay valvas desarticuladas y algunas veces incompletas de un solo individuo de los géneros Posidoniella, Septimyalina, Bakevellia o Leptodesma, que incluyen formas epifaunales o semi-infaunales que comúnmente vivían en aguas salobres; y de Edmondia o Cardiomorpha que fueron infaunales y cuyo registro en otros trabajos se asocia a depósitos de aguas poco profundas (Girty, 1910a; Ciriacks, 1963; Runnegar y Newell, 1974; Dickins, 1999).
El hecho de que los bivalvos infaunales sean abundantes en las concreciones y que presenten las valvas articuladas y, por otra parte, que los bivalvos epifaunales estén desarticulados y sus valvas muestren escaso desgaste, es congruente con los hábitos de vida de los géneros estudiados y sugiere cambios en las condiciones de depósito. Los primeros representan elementos autóctonos o parauctóctonos, mientras que los segundos que vivían a menor profundidad, como los bivalvos Posidoniella sp, especie epifaunal cuyo registro en el campo presenta valvas grandes completas, apoya la idea de un medio de baja energía y transporte moderado o rápido. Los restos vegetales asociados se interpretan como elementos alóctonos al ser transportados desde el continente hasta el mar mediante corrientes fluviales.
De acuerdo con los elementos taxonómicos y tafonómicos, y con la lutita fosilífera, se interpreta un ambiente de plataforma de aguas poco profundas de baja energía, influenciado por la desembocadura de ríos y el transporte de bivalvos provenientes de estuarios, lo cual es congruente con el modelo de Juárez-Arriaga (2006).
6. Consideraciones biogeográficas
Las reconstrucciones paleogeográficas coinciden en colocar el norte del territorio mexicano como parte de Laurusia, en una región de latitudes bajas y formando parte de un extenso mar somero (Bambach et al., 1980; Ziegler et al., 1997). Durante el Carbonífero, este mar somero tenía continuidad hacia el norte hasta la latitud más alta de América del Norte, y hacia el sur hasta Gondwana, y sólo era interrumpido por algunos terrenos poco elevados. Sin embargo, como resultado de la formación de Pangea, su extensión se redujo durante el Pérmico medio. En este mar quedó comprendida el área de estudio.
Los géneros y especies referidos presentan afinidad con faunas del Pérmico medio de América del Norte que se han relacionado con las provincias Cordillerana y Laurencia Sur del Reino Americano de acuerdo con el modelo de Bambach (1990) y con las provincias Cordillerana y Grandiana propuestas por Yancey (1975), como parte del reino Paleoecuatorial (Shen et al., 2009) (Figura 6).
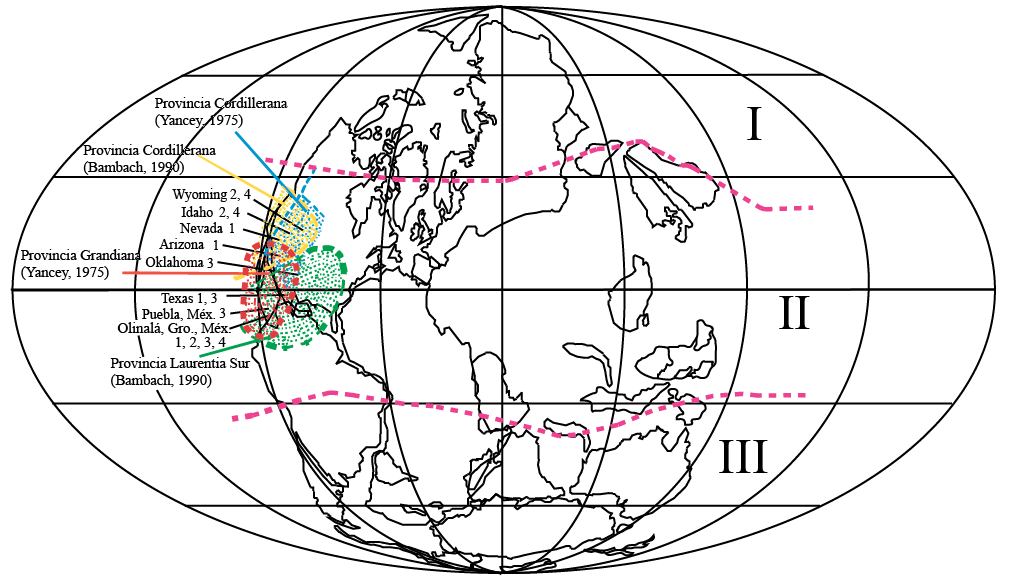
Figura 6. Distribución paleogeográfica de las especies estudiadas, durante el Pérmico medio. Los números indican las especies: 1. Nuculavus levatiformis, 2. Palaeoneilo mcchesneyana, 3. Septimyalina burmai, 4. Edmondia phosphatica. Las áreas en las que se encuentra el registro de bivalvos corresponden a mares poco profundos. Reinos biogeográficos: I. Boreal; II. Paleoecuatorial y III. Gondwánico y extensión de la Provincia Grandiana (Yancey, 1975) de acuerdo a Shen et al., 2009). Mapa base modificado de Ziegler et al., 1997.
7. Paleontología sistemática
En la clasificación siguiente se adoptan la nomenclatura y clasificación propuestos por Bieler et al. (2010). Para las categorías inferiores a familia se siguen los criterios de Newell y LaRocque en Cox et al. (1969). El material descrito se encuentra depositado en la colección del Museo de Paleontología de la Facultad de Ciencias, Universidad Nacional Autónoma de México con el acrónimo FCMP.
Clase Bivalvia Linnaeus, 1758
Orden Nuculida Dall, 1889
Superfamilia Nuculoidea Gray, 1824
Familia Nuculidae Gray, 1824
Género Nuculavus Chernyshev, 1947
Especie tipo. Nuculavus minuta Chernyshev, 1947, por designación original. Carbonífero de Rusia.
Diagnosis. Nucúlidos pequeños, trigonales, con ornamentación concéntrica que consiste en líneas de crecimiento finas e irregulares, umbo ligeramente desplazado hacia la parte posterior, resilífero pequeño, abierto, poco profundo y triangular. Interior del margen ventral liso (Yancey, 1978).
Nuculavus levatiformis (Walcott, 1884)
Figuras 7.1-7.3
Nucula levatiforme Walcott, 1884, p. 241, lám. 22, fig. 1, 1a.
Nucula levatiformis Walcott. Girty in Lee y Girty 1909, p. 74, lám. 10, figs. 7, 8; Clifton, 1942, p. 693.
Palaeonucula levatiformis (Walcott) 1884. Chronic, 1952, p. 138, lám. 6, figs. 4-9; Winters, 1963, p. 49, lám. 6, figs. 4-7a; Mayou, 1967, p. 118, lám. 1, fig. 10; Yancey, 1969, p. 6, figs. 18-20.
Palaeonucula cf. levatiformis (Walcott) 1884. Chronic en Newell et al., 1953, p. 151, lám. 33, figs. 2a, b.
Nuculavus levatiformis (Walcott) 1884. Yancey, 1978, p. 316, lám. 7, figs. 6-10; Sterren, 2000, p. 428, figs. 9.A, 9.B; Anelli et al., 2002, p. 168, figs. 3c-d, 4; Sterren, 2004, p. 60, figs. 3, A-B.
Descripción. Concha pequeña, equivalva, inequilateral y subtrigonal. Dentición incompleta, con los dientes anteriores y posteriores de tamaño uniforme. Margen ventral redondeado. Carena tenue en el borde anterior. Superficie con estrías concéntricas finas y numerosas. Interior con impresiones de los músculos aductores isomiarios; también se observa la línea palial.
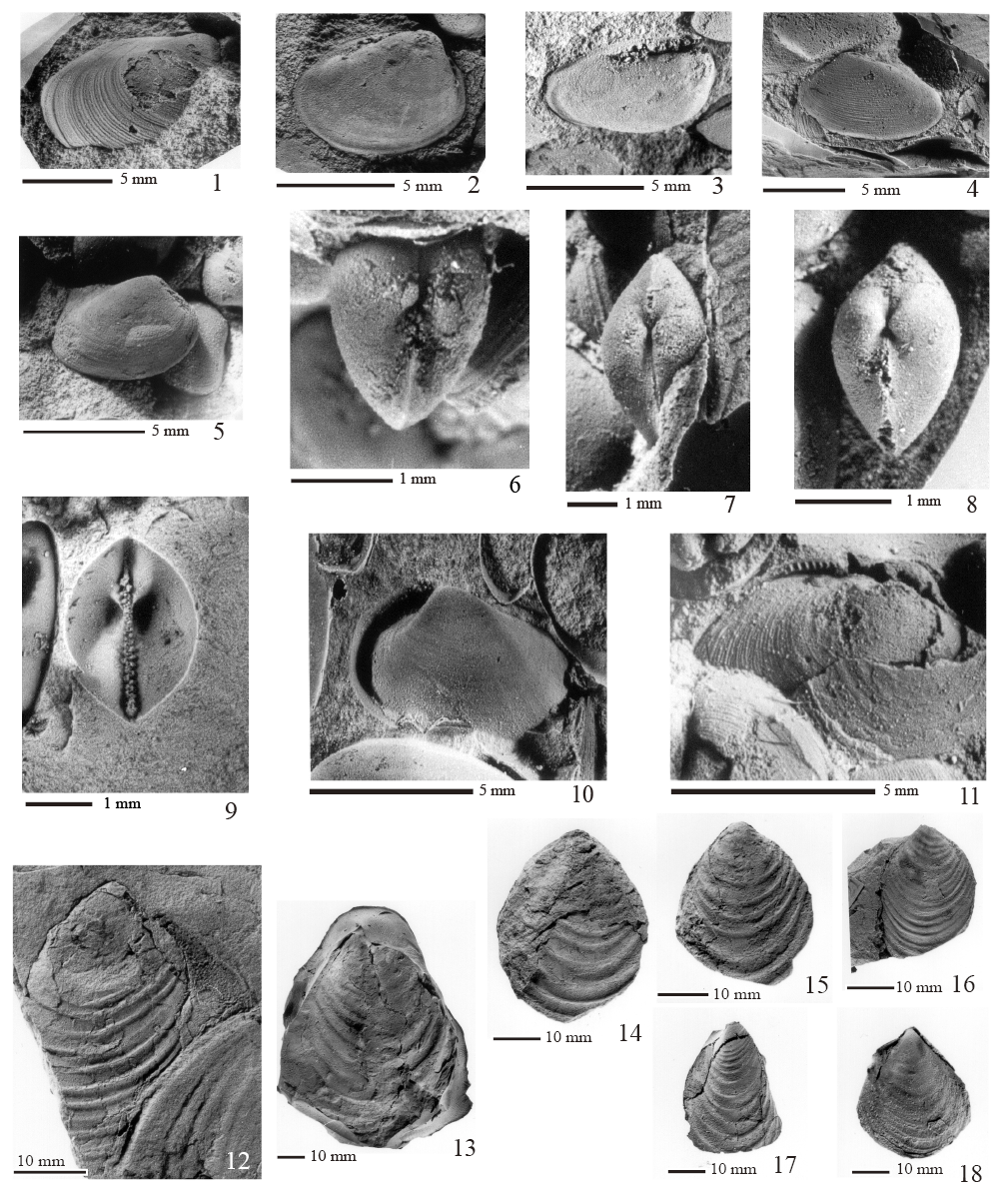
Material examinado. Un molde externo e interno de una valva izquierda (FCMP 841) y un molde interno de ambas valvas articuladas (FCMP 842).
Medidas. (Tabla 1)
Distribución. Nuculavus levatiformis se ha referido para el Pérmico Inferior-Medio (Asseliano-Wordiano) de Arizona, Texas, Nevada y Utah en los Estados Unidos (Clifton, 1942; Chronic, 1952; Winters, 1963; Cooper y Grant, 1972; Yancey, 1978) y en el Carbonífero Superior-Pérmico Inferior (Gzheliano–Asseliano) de Argentina (Sterren, 2000; 2004). Chronic en Newell et al. (1953) la refiere para el Paleozoico Superior (Asseliano-Artinskiano) de Perú y Anelli et al. (2002) refieren esta especie para el Pensilvánico Medio de la cuenca de Parnaíba en Brasil.
Discusión. Nuculavus se diferencia de otros géneros similares: Nuculopsis presenta valvas de forma subcuadrada y alargada (Yancey, 1978 en Anelli et al., 2002); Nucula tiene el interior del margen ventral crenulado y un resilífero que interrumpe la continuidad de la dentición (Elías, 1957 en Anelli et al., 2002).
Yancey (1978) distingue a Nuculavus levatiformis de otras especies del género Nuculavus por presentar un umbo menos prominente, valvas moderadamente convexas y márgenes anterior y posterior redondeados. El mismo autor reconoce una amplia variabilidad y diferencias regionales dentro de la especie.
Los ejemplares estudiados presentan características afines a los ejemplares mostrados por Yancey (1978) y por Anelli et al. (2002). En los ejemplares estudiados no se observa el pequeño resilífero triangular, sin embargo sí se presentan los demás caracteres de la especie.
Localidad. Camino Olinalá-El Progreso, localidad 2.
Orden Nuculanida Carter, Campbell y Campbell, 2000
Superfamilia Nuculanoidea Adams y Adams, 1858
Familia Mallettidae Adams y Adams, 1858
Género Palaeoneilo Hall y Withfield, 186
Especie tipo. Nuculites constricta Conrad, 1842, por designación subsecuente de Hall, 1885. Devónico Medio de Nueva York.
Diagnosis. Valvas posteriormente alargadas, con o sin resilífero, usualmente con seno palial. Muesca radial posterior apenas visible, ornamentación concéntrica, sin septo interno (McAlester, 1969).
Palaeoneilo mcchesneyana (Girty, 1910a)
Figuras 7.4-7.11
Yoldia mcchesneyana Girty, 1910a, p. 39, lám. 4, figs. 4-6.
Nucula mcchesneyana Branson, 1948, p. 636.
Anthraconeilo mcchesneyana Girty, 1910 a. Yancey, 1978, p. 324, lám. 8, figs. 13-15.
Palaeoneilo mcchesneyana (Girty). Ciriacks, 1963, p. 37, lám. 4, figs. 12-16; Fang, 1987, p. 364, lám. 1, figs. 8-10.
Descripción. Concha equivalva, con valvas inequilaterales, alargadas e infladas; umbo poco prominente y ortogiro desplazado hacia el centro entre los márgenes anterodorsal y posterodorsal. Margen posterodorsal más grande que el anterodorsal; margen ventral redondeado. Superficie con ornamentación comarginal consistente en líneas delgadas poco espaciadas entre sí.
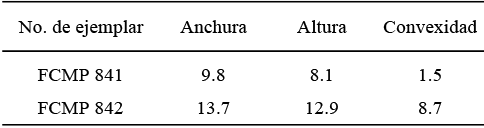
Material examinado. Un molde externo de una valva derecha (FCMP 843), un molde compuesto de una valva izquierda (FCMP 844) y seis ejemplares articulados (FCMP 845-848, FCMP 849 y FCMP 850), de los cuales FCMP 849 tiene las valvas abiertas en forma de mariposa. Además se presentan varios moldes externos encontrados en el interior de concreciones de lutita Figuras 5.1 y 5.2 cuya preservación no permite medir sus dimensiones. Se trata de la segunda especie de bivalvo más abundante en las localidades estudiadas.
Medidas. (Tabla 2)
Distribución. Palaeoneilo mcchesneyana ha sido reportada para los estados de Idaho, Wyoming (Girty, 1910a; Ciriacks, 1963) y Nevada (Yancey, 1978), en Estados Unidos, y en Hunan, China (Fang, 1987). Su alcance estratigráfico abarca desde el Artinskiano hasta el Wuchiapingiano.
Discusión. Palaeoneilo mcchesneyana se diferencia de P. sera Girty (1910b) por tener el margen totalmente redondeado y no sinuoso hacia su parte posterior formando una ligera muesca, y su umbo es menos prominente (Hoare et al., 1989; Hoare, 1993); de P. tebagaensis (Termier y Termier 1959 en Boyd y Newell, 1979) por sus umbos ortogiros que no están aplanados; de P. oweni (McChesney en Hoare et al., 1979) por poseer valvas con márgenes posterodorsal y posteroventral que se juntan sin formar un contorno agudo; de P. qinzhouensis (Weihong et al., 2007) por presentar una charnela no arqueada y umbo ortogiro.
Localidad. Camino Olinalá-El Progreso, localidad 2.
Tabla 2. Dimensiones de Palaeoneilo mcchesneyana (en mm).
Orden Pteriida Newell, 1965
Superfamilia Ambonychioidea Miller, 1877
Familia Myalinidae Frech, 1891
Género Posidoniella De Koninck, 1885
Especie tipo. Inoceramus vetustus Sowerby, 1829, por designación original. Carbonífero Inferior de Inglaterra.
Diagnosis. Conchas ventricosas con el aspecto de los inocerámidos, sin ornamentación radial, con aurícula anterior. Con una delgada capa prismática (Newell en Cox et al., 1969).
Posidoniella sp.
Figuras 7.12-7.18, y Figuras 9.1, 9.2
Descripción. Valvas adultas suborbiculares, con umbo ortogiro próximo a la parte posterior que sobresale del margen dorsal. Pequeña prolongación a manera de aurícula con posición anterior. La ornamentación consiste en pliegues concéntricos gruesos y ampliamente espaciados entre sí; carecen de ornamentación radial.
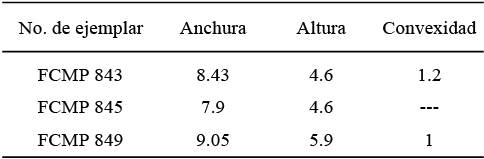
Material examinado. Once moldes externos de valvas izquierdas (FCMP 851-861) y nueve de valvas derechas (FCMP 862-870), dentro de los cuales, los que presentan mejor conservación son: FCMP 851-856 y FCMP 862-864.
Medidas. (Tabla 3)
Distribución. Posidoniella ha sido referido para el Carbonífero (Viseano-Namuriano), con reportes en el Pensilvánico (Cox et al., 1969). También fue reportado para el Carbonífero Inferior de Chile por Isaacson y Dutro (1999) en el miembro superior de la Formación Las Zorritas. Para el Misisípico Superior del Reino Unido (Wolfenden, 1958) se mencionaron las especies P. gibbosa Hind 1897 y P. vetusta (Sowerby, 1829). En España se refirió para el Namuriano-Westphaliano (Sampelayo y Sampelayo, 1947). González-Arreola et al. (1994) lo refirieron para la Formación Olinalá asociado a braquiópodos (Cancrinella rugosa Cloud, 1944, Thamnosia depressa (Cooper, 1953) y Orbiculoidea ovalis Cloud, 1944), gasterópodos (Babylonites carinatus Yochelson, 1956), amonoideos (Stacheoceras toumanskyae Miller y Furnish, 1940, Paraceltites elegans Girty, 1908 y Waagenoceras dieneri (Böse, 1919), y el conulárido Paraconularia sp., que en conjunto indican una edad Pérmica. Quiroz-Barroso et al. (2012), refirieron este género para el Cisuraliano (Pérmico inferior) de Otlamalacatla, Hidalgo.
Discusión. Los ejemplares de esta especie son los más abundantes en el área, aunque su preservación no es la mejor debido a que los moldes externos se encuentran sobre láminas finas de lutita gris clara a oscura, que se rompen fácilmente al recolectar los ejemplares. El material ilustrado por González-Arreola et al. (1994) y el que se recolectó para este estudio es incompleto, pues solamente se tienen las partes próximas al umbo (Figura 8).
Debido a la pobre preservación de los ejemplares se deja abierta la determinación.
Localidad. Camino Olinalá-El Progreso, localidades 1 y 2.

Figura 8. 1, Ejemplar de Posidoniella sp. observado en el afloramiento; 2, reconstrucción del mismo ejemplar, las líneas punteadas representan las partes más frágiles de la valva que se quiebran al recolectarlas.
Tabla 3. Dimensiones de Posidoniella sp. (en mm).
Género Septimyalina Newell, 1942
Especie tipo. Myalina perattenuata Meek y Hayden, 1858, por designación original. Pensilvánico Superior de Kansas, Estados Unidos.
Diagnosis. Concha prosoclina, romboidal, falcada o aclina; delgada, con umbos extendidos y delgados, con lóbulo anterior obsoleto debajo de cada umbo; ambas valvas lisas o rugosas; inequivalvas; cavidad umbonal anterodorsalmente cubierta por una placa charnelar ancha; dentición débil, como en Myalina (Newell en Cox et al., 1969).
Septimyalina burmai (Newell, 1942)
Figuras 9.3-9.8
?Myalina apachesi Marcou, 1858, p. 44, lám. 7, figs. 6, 6a.
Myalina apachesi Marcou. Lee y Girty, 1909, p. 81, lám. 9, figs. 6, 7; Clifton, 1942, p. 690, lám. 101, figs. 17, 18; lám. 102, figs. 3, 4.
Septimyalina burmai, Newell, 1942, p. 67, lám. 12, figs. 1-6; Mudge et al., 1962, lám. 15, fig. 21; Kues, 1995, lám. 7, figs. 30-34.
Descripción. Concha mediana, mitiliforme, equivalva y prosoclina; umbo prosogiro y puntiagudo, poco prominente; margen cardinal recto, formando un ángulo obtuso con el margen posterior, que es ligeramente convexo; margen anterior convexo en la parte superior, recto en la inferior; margen ventral redondeado. Ambas valvas marcadas con laminillas concéntricas espaciadas regularmente en forma de arrugas, sin ornamentación radial.
Material examinado. Siete moldes externos de valvas derechas (FCMP 871-877), de los que el ejemplar FCMP 871 es una forma juvenil; un molde externo de una valva izquierda (FCMP 878); y dos moldes externos con las valvas articuladas (FCMP 879 y FCMP 880), de los cuales la última está abierta en forma de mariposa.
Medidas. (Tabla 4)
Distribución. Septimyalina burmai ha sido referida para el Gzheliano-Kunguriano de América del Norte. Ha sido referida para América del Norte: en Puebla, México (Sour-Tovar et al., 2000) y los estados de Kansas (Newell, 1942; Mudge et al., 1962), Nuevo México (Kues, 1995) y Oklahoma (Clifton, 1942) en Estados Unidos. En Sudamérica ha sido referido para Brasil (Anelli et al., 2006) y Perú (Newell et al., 1953).
Discusión. De acuerdo con Newell (1942), no es posible comprobar la sinonimia con el material estudiado por Marcou (1858), pues los ejemplares en que se basa el segundo trabajo se perdieron y se desconocen tanto la localidad de recolección como el horizonte exacto.
S. perattenuata (Meek y Hayden, 1858) presenta la marca del ligamento en la charnela, conformada por una región muy amplia en la concha que muestra una serie de líneas donde se inserta el ligamento. En el ejemplar estudiado, la charnela parece no ser tan grande como para que exista tal área. En el ejemplar de Newell (1942) se observa que la parte de la charnela que tiene la inserción del ligamento es muy ancha y esta área no aparece en el ejemplar estudiado. Hoare et al. (1979) incluye dentro de esta especie ejemplares con una variabilidad evidente, desde una concha muy alargada y pequeña y carente de una aurícula, hasta conchas grandes, alargadas y anchas, con una aurícula grande.
Anelli et al. (2006) presentaron varios moldes internos y un externo de S. perattenuata, de los que, en el último, se observa una notable diferencia con S. burmai: la presencia de un plegamiento en la parte antero-ventral de la valva izquierda. Dicho plegamiento no se presenta en S. burmai
Septimyalina burmai se diferencia de S. orthonota (Mather, 1915) por el umbo puntiagudo y prominente; de S. orbiculata Newell, 1942 por su charnela menos amplia, su concha menos inequilateral y su umbo menos largo; de S. sinuosa (Morningstar, 1922) por su concha menos ancha, un umbo mucho menos puntiagudo y una charnela menos amplia. S. perattenuata, S. orthonota y S. orbiculata pertenecen al Pensilvánico, y esta última pudo ser ancestro de S. scitula Newell, 1942, sin relación con S. burmai, mientras que S. burmai, S. perattenuata y S. orthonota conforman un género natural, pues de acuerdo con Newell (1942) aparecen sucesivamente en la columna geológica.
Las especies restantes del género son lo suficientemente distintas a S. burmai pues es fácil distinguir sus diferencias. Por lo tanto, con base en la comparación de los ejemplares estudiados con otras especies de este género, se determina que pertenece a Septimyalina burmai debido a que carece de aurículas, no tiene una concha ancha y sus umbos son puntiagudos y ligeramente curvados.
Localidad. Camino Olinalá-El Progreso, localidades 1 y 2.
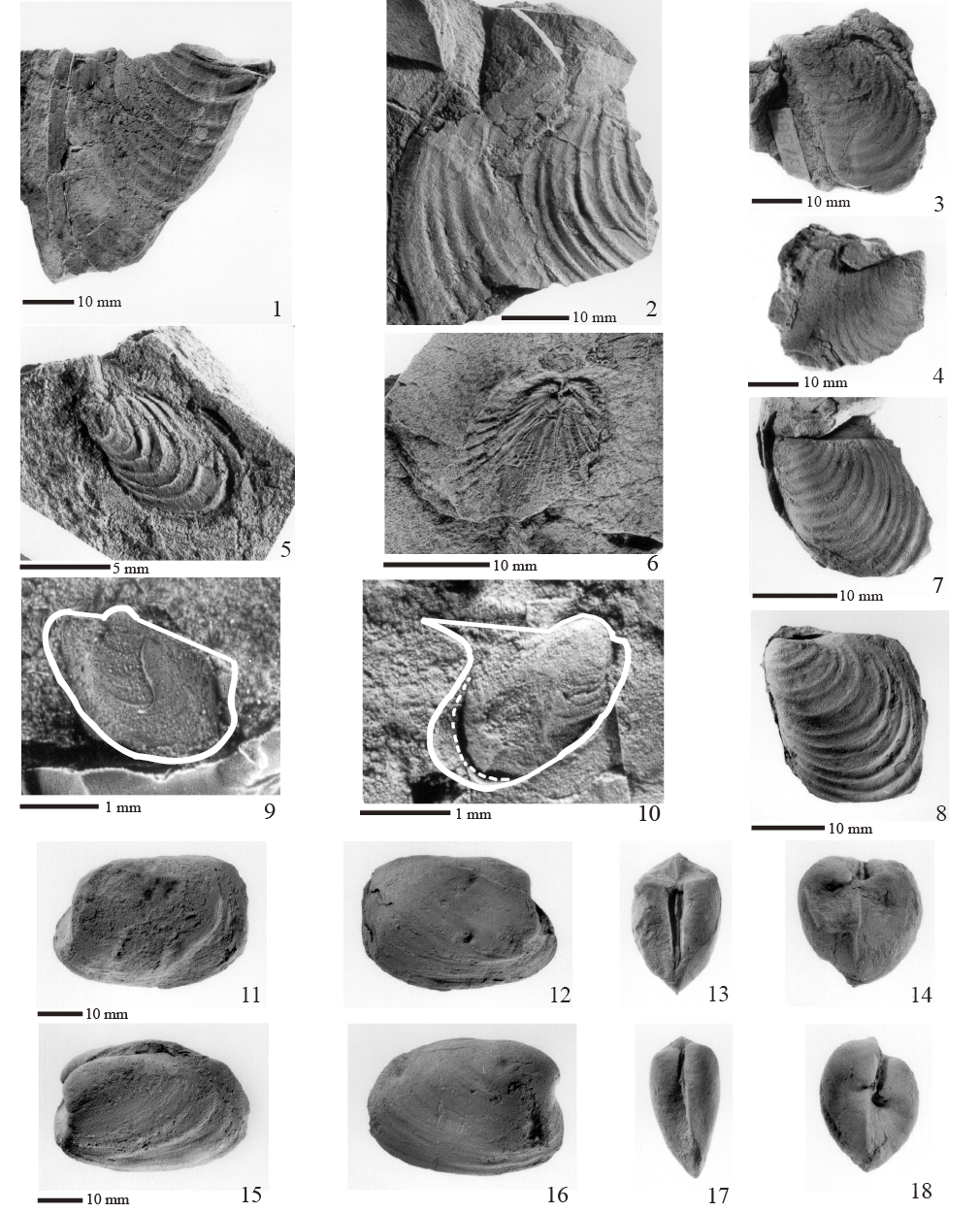
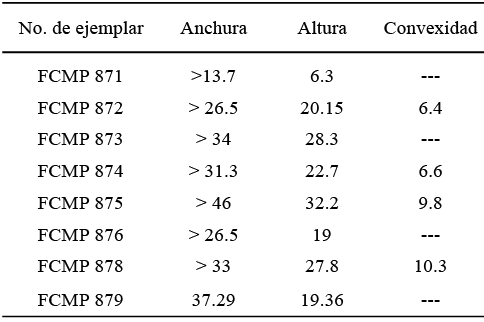
Figura 9. 1, 2: Posidoniella sp.; 1, fragmento del margen ventral, ejemplar FCMP 864; 2, fragmentos de dos valvas, ejemplar FCMP 852; 3-8, Septimyalina burmai (Newell, 1942); 3,4, moldes externos de valvas izquierda y derecha, articuladas, abiertas en forma de mariposa, ejemplar FCMP 879; 5, molde externo de la valva izquierda, juvenil, ejemplar FCMP 871; 6, molde externo, vista anterior de ambas valvas articuladas, abiertas y deformadas, ejemplar FCMP 880; 7, molde externo de la valva izquierda, ejemplar FCMP 872; 8, molde externo de la valva izquierda, ejemplar FCMP 873; 9, Bakevellia sp. Molde externo de la valva izquierda, ejemplar FCMP 881; 10, Leptodesma (Leptodesma) sp. Molde compuesto imperfecto de la valva derecha, ejemplar FCMP 882; 11-18, Edmondia phosphatica Girty, 1910a; 11, valva izquierda, 12, valva derecha, 13, vista dorsal, 14, vista anterior, ejemplar FCMP 883; 15, valva izquierda, 16, valva derecha, 17, vista dorsal, 18, vista anterior, ejemplar FCMP 885.
Superfamilia Pterioidea Gray, 1847
Familia Bakevelliidae King, 1850
Género Bakevellia King, 1848
Especie tipo. Avicula antiqua von Münster en Goldfuss, 1836, por designación original. Pérmico Superior de Inglaterra.
Diagnosis. Concha de pequeña a mediana, de forma trapezoidal o romboidal, no muy alargada; la aurícula anterior puede ser de pequeña a moderadamente grande, el ala posterior es obtusa o muy puntiaguda; las valvas nunca son extremadamente inequivalvas; con apertura anterior estrecha. Pérmico-Cretácico, cosmopolita. (Cox et al., 1969).
Bakevellia sp.
Figura 9.9
Descripción. Valva pequeña, subcuadrada. Umbo pequeño. La aurícula anterior es angosta y puntiaguda; la aurícula posterior es amplia, de ángulo obtuso y más grande que la aurícula anterior. Ornamentación no visible.
Material examinado. Un molde imperfecto de una valva izquierda (FCMP 881).
Medidas. 3.5 mm de anchura y 2.3 mm de altura.
Distribución. El alcance estratigráfico del género Bakevellia va desde el Serpukhoviano (Pérmico) hasta el Maastrichtiano (Cretácico). Para el Pérmico, el género Bakevellia tiene reportes provenientes de Canadá (Harker y Thorsteinsson, 1960), Italia (Posenato et al., 2005), México (Blodgett et al., 2002), Reino Unido (Logan, 1967) y en Estados Unidos es donde existe una mayor cantidad de referencias provenientes de Arizona (Chronic, 1952), Idaho, Texas y Wyoming (Ciriacks, 1963), Nevada (Yancey y Stevens, 1981) y Nuevo México (Kues, 1995).
Discusión. Kues (1995) presentó numerosas conchas halladas en calizas, y las determinó como Bakevellia sulcata Geinitz (1866). En dicho trabajo se ilustran valvas izquierdas similares al ejemplar que aquí se determina; Kues (1995) indicó que se trata de valvas de tamaños menores a 5 mm. No es posible comparar la valva que aquí se determina con la de Kues, pues aunque describe a detalle sus ejemplares, sus fotografías no permiten observar bien la morfología; Winters (1963) reportó especímenes de B. sulcata cuya área ventral es mucho más larga que el ejemplar que aquí se determina, además no es posible la comparación con esta especie porque la valva que aquí se describe no presenta el sulcus, un carácter diagnóstico para la especie.
Localidad. Camino Olinalá-El Progreso, localidad 1.
Familia Pterineidae Meek, 1864
Género Leptodesma Hall, 1883
Especie tipo. Leptodesma potens Hall, 1883, por designación subsecuente de Miller, 1889. Devónico Superior de Nueva York, Estados Unidos.
Diagnosis. Sin ornamentación radial, umbo izquierdo más prominente que el derecho; ala posterior comúnmente bien desarrollada; contrafuerte miofórico débil; pocos dientes cardinales y variables en número (Newell y LaRocque en Cox et al., 1969).
Subgénero Leptodesma Hall, 1883
Leptodesma (Leptodesma) sp.
Figura 9.10
Descripción. Concha pequeña, inequivalva y poco convexa. Margen anterior ligeramente redondeado; margen posterior paralelo al anterior, convexo hacia el margen ventral; margen cardinal recto y amplio, mayor que la longitud de la concha. Umbo pequeño y poco prominente. Aurícula anterior muy reducida; aurícula posterior en forma de ala muy aguda que termina en forma de espina. Superficie ornamentada con líneas comarginales finas y espaciadas.
Material examinado. Un ejemplar (FCMP 882) que consiste en la impresión externa de la valva izquierda casi totalmente cubierta por el molde compuesto imperfecto de la valva derecha. Las valvas están desplazadas.
Medidas. 4.2 mm de anchura y 3.4 mm de altura.
Distribución. El alcance estratigráfico del subgénero Leptodesma (Leptodesma) comprende desde el Telychiano (Silúrico Inferior) al Changhsingiano (Pérmico Superior). Es cosmopolita.
Discusión. Se asigna a Leptodesma (Leptodesma) por poseer una aurícula anterior angular, que lo distingue del otro subgénero, Leptodesma (Leiopteria) Hall, 1883, en el cual es lobulada (Newell y La Rocque en Cox et al., 1969). El estado precario del único ejemplar no permite conocer características diagnósticas, por lo que se deja abierta la asignación de especie.
Localidad. Camino Olinalá-El Progreso, localidad 1.
Orden Pholadomyida Newell, 1965 [Anomalodesmata Dall, 1889]
Superfamilia Edmondioidea King, 1850
Familia Edmondiidae King, 1850
Género Edmondia De Koninck, 1841
Especie tipo. Isocardia unioniformis Phillips, 1836, por designación original. Carbonífero Inferior de Irlanda.
Diagnosis. Concha ovoide a elíptica alargada, regularmente gibosa. Sin carenas laterales o sulco; márgenes cerrados; umbos pequeños, prosogiros, situados de un cuarto a un tercio detrás del margen anterior; las valvas generalmente están ornamentadas con algunas líneas concéntricas irregulares o líneas de crecimiento; placa charnelar internamente reforzada por una cresta casi paralela (Newell en Cox et al., 1969).
Edmondia phosphatica Girty, 1910a
Figuras 9.11-9.18
Edmondia? phosphatica Girty, 1910a, p. 36, lám. 6, fig. 9.
Edmondia phosphatica Girty. Ciriacks, 1963, p. 74, lám. 11, figs. 12-15.
Descripción. Concha subcuadrada, muy inflada, inequilateral. Umbos prosogiros, muy próximos a la parte anterior. El margen anterodorsal presenta un truncamiento, mientras que el posterodorsal es largo y ocupa la mayor parte de la zona dorsal. Márgenes dorsal y ventral planos, más o menos paralelos entre sí. Charnela larga. La ornamentación es comarginal y consiste en pliegues gruesos muy pronunciados.
Material examinado. Tres moldes externos de valvas articuladas (FCMP 883, FCMP 884 y FCMP 885).
Medidas. (Tabla 5).
Distribución. Edmondia phosphatica se ha descrito para el Roadiano de Idaho y Wyoming (Girty, 1910a; Ciriacks, 1963), en Estados Unidos.
Discusión. El género Edmondia se distingue por tener una placa charnelar, que en moldes internos se observa como un surco paralelo al margen dorsal. La cara ventral de esta placa presenta un conjunto de inserciones musculares circulares, y no debe interpretarse como el sitio de fijación de un ligamento interno (Runnegar y Newell, 1974).
Edmondia phosphatica se diferencia de E. nodulifera (Hoare et al., 1978) por presentar una ornamentación con líneas gruesas; de E.? bellula Girty, 1908 por su umbo prosogiro, muy próximo a la parte anterior.
Ciriacks (1963) menciona que Edmondia phosphatica presenta valvas con ornamentación delgada; sin embargo, en la lámina 11, figuras 12-15 de dicho trabajo se observa una ornamentación más gruesa, muy similar a la que presentan los ejemplares de México. Se determina que se trata de esta especie porque los caracteres observados en el trabajo de Ciriacks coinciden con los de los ejemplares descritos aquí. También se puede observar que los ejemplares estudiados aquí tienen unos márgenes dorsal y ventral lisos y casi paralelos entre sí, lo cual concuerda con los ejemplares de Ciriacks.
Localidad. Camino Olinalá-El Progreso, localidad 2.
|
BOLETÍN DE LA SOCIEDAD GEOLÓGICA MEXICANA Vol 65, Núm. 3, 2013, P. 533-551 |
|
Bivalvos guadalupianos (Pérmico medio) de la Formación Olinalá, Guerrero, México
Miguel Guerrero-Sánchez1, Sara A. Quiroz-Barroso2,*
1Facultad de Ciencias, Universidad Nacional Autónoma de México, Ciudad Universitaria, 04510 México D.F., México.
2Museo de Paleontología, Departamento de Biología Evolutiva, Facultad de Ciencias, Universidad Nacional Autónoma de México, Ciudad Universitaria, 04510 México D.F., México.
* This email address is being protected from spambots. You need JavaScript enabled to view it.
Resumen
El material estudiado proviene de rocas que afloran al noreste del poblado Olinalá, en el estado de Guerrero. Las facies marinas de la Formación Olinalá están incluidas en lutita gris claro a oscuro con fósiles de braquiópodos, gasterópodos, bivalvos, cefalópodos, tallos de crinoideos, madera y otros restos de plantas, localmente alojados en concreciones. Entre la diversidad de los bivalvos se encuentran las especies infaunales Nuculavus levatiformis, Palaeoneilo mcchesneyana, Edmondia phosphatica y ?Cardiomorpha sp.; las especies epifaunales que vivían adheridas a diferentes sustratos Posidoniella sp., Septimyalina burmai y Leptodesma (Leptodesma) sp.; así como la especie semi-infaunal Bakevellia sp. Los hábitos de vida de los bivalvos y las características de su preservación indican que fueron poco transportados antes de su depósito final, lo cual ocurrió en una plataforma de aguas poco profundas. Las especies de bivalvos estudiadas concuerdan con la edad pérmica de esta formación, establecida previamente por otros autores con base en el estudio de amonites y foraminíferos, y apoyan la idea de su relación con faunas contemporáneas de América del Norte.
Palabras clave: Formación Olinalá, bivalvos, Pérmico medio, México.
Abstract
The study material comes from rocks that outcrop northeast of the town Olinalá, in Guerrero state. The marine facies of Olinalá Formation are included in light to dark gray shale with fossils of brachiopods, gastropods, bivalves, cephalopods, crinoids stems, wood and other plant remains, locally included in concretions. The diversity of bivalves is represented by the infaunal species Nuculavus levatiformis, Palaeoneilo mcchesneyana, Edmondia phosphatica, and ?Cardiomorpha sp.; the epifaunal species, adhered to different substrates Posidoniella sp., Septimyalina burmai, and Leptodesma (Leptodesma) sp.; and the semi-infaunal species Bakevellia sp. The lifestyle and preservation characteristics of bivalves indicate that they were shortly transported before its final deposition, which occurred on a platform, in shallow water. The studied bivalve species are consistent with the permian age of this formation, considered previously by other authors based on the study of ammonites and foraminifera, and support the idea of their relationship with contemporary faunas of North America.
Palabras clave: Olinalá Formation, bivalves, middle Permian, Mexico.
1. Introducción
Las rocas que afloran en la parte noreste del poblado Olinalá, en el estado de Guerrero, constituyen una sucesión de más de 800 m de espesor que ha sido denominada como Formación Los Arcos (Corona-Esquivel, 1981 [1983]) y Formación Olinalá (Flores de Dios y Buitrón, 1982), siendo este último nombre el reconocido como válido en el Léxico Estratigráfico del Servicio Geológico Mexicano (SGM, 2006).
La Formación Olinalá está constituida por una diversidad de facies terrígenas (marinas y continentales) y carbonatadas (marinas), cuyos ambientes de depósito varían de fluviales a margen de rampa, costeros y plataforma externa (Juárez-Arriaga, 2006). De ellas, las facies marinas de lutita son las más abundantes tanto en la base como en la cima de la formación y consisten en lutita gris claro a oscuro finamente laminada, con alto contenido de biotita en la base y abundantes concreciones, generalmente paralelas a la estratificación (Juárez-Arriaga, 2006). En cuanto a su fauna se ha mencionado la presencia de calcimicrobios (Tubiphytes sp.), foraminíferos, briozoarios, braquiópodos, bivalvos, gasterópodos, amonoideos, artrópodos (ostrácodos y trilobites), crinoideos, así como restos vegetales (Corona-Esquivel, 1981 [1983], 1985; Flores de Dios y Buitrón, 1982; Flores de Dios, 1986; González-Arreola et al., 1994; Silva-Pineda et al., 1998; Flores de Dios et al., 2000; Garibay-Romero, 2000; Esquivel-Macías, 2000; Esquivel-Macías et al., 2004; Vachard et al., 2004; Juárez-Arriaga, 2006).
La presencia de bivalvos en la facies de lutita marina se menciona e ilustra en los estudios de González-Arreola et al. (1994) y Juárez-Arriaga (2006). En el presente trabajo se describen las especies de bivalvos de la lutita media de esta formación y se propone una interpretación acerca de su presencia en rocas del Paleozoico superior de México, analizándose sus hábitos de vida, así como su distribución geográfica y estratigráfica.
2. Ubicación del área de estudio
El material estudiado se recolectó en rocas que afloran al noreste de Olinalá, en el estado de Guerrero (Figura 1).
Localidad 1. Sobre el camino de terracería que parte desde Olinalá hacia el poblado El Progreso, con coordenadas 17° 46.864’ latitud N y 98° 43.761’ longitud W. Lutita de color gris oscuro que intemperiza en forma de láminas delgadas, con manchas color ocre debidas a la oxidación y con contenido fósil. Contiene concreciones que varían en tamaño de 1.5 a 20 cm, en muchas de las cuales hay contenido fósil; algunas de las concreciones son alargadas y se encuentran sobre el plano de estratificación. Se presentan moldes de ejemplares grandes del bivalvo Posidoniella De Koninck, 1885, así como moldes de otros bivalvos (Septimyalina Newell, 1942; Bakevellia King, 1848 y Leptodesma Hall, 1883), gasterópodos, amonoideos y un fragmento de un tallo de crinoideo. Se observan impresiones de restos vegetales.
Localidad 2. Sobre el camino de terracería que parte desde Olinalá hacia el poblado El Progreso, con coordenadas 17° 47.160’ latitud N y 98° 43.861’ longitud W. Lutita negra con estratificación paralela. El echado de las capas es de casi 90°. Están presentes moldes de braquiópodos, bivalvos (Nuculavus Chernyshev, 1947; Palaeoneilo Hall y Withfield, 1869; Posidoniella, Septimyalina, Edmondia De Koninck, 1841 y ?Cardiomorpha De Koninck, 1844), gasterópodos, nautiloideos y placas de crinoideos.
Figura 1. Área de estudio. Las barras negras con números representan la ubicación de las localidades de recolecta
3. Estratigrafía
Las rocas de la Formación Olinalá yacen discordantemente sobre rocas metamórficas del Complejo Acatlán de edad Ordovícico Tardío-Devónico Tardío (Ortega-Gutiérrez et al., 1999) y están cubiertas discordantemente por la Ignimbrita Las Lluvias de edad Triásico?-Jurásico Medio (Corona-Esquivel, 1981 [1983]; García-Díaz et al., 2000). A su vez, esta unidad está cubierta por el Conglomerado Cualac y el Grupo Tecocoyunca (Erben, 1956) del Jurásico Medio.
Corona-Esquivel (1981 [1983]), reconoció siete unidades en la Formación Olinalá. La unidad uno es la base de la formación y es un conglomerado poligenético conformado por guijarros sub-redondeados de esquisto de mica, cuarzo blanco, cuarcita, esquisto verde de clorita y algunos fragmentos de filita. La unidad dos consiste en limolita negra y lutita gris oscura a negra, con algunas capas de arenisca gris de grano fino, de 15 cm de espesor y abundantes concreciones limolíticas; en la base se presenta un horizonte con amonites y bivalvos dentro de las concreciones. La unidad tres se divide en dos partes: a) una inferior, que consiste en capas gruesas de conglomerado poligenético con guijarros de gneis sub-angulosos y redondeados, esquisto, pizarra negra y cuarcita gris claro; y b) una superior con arenisca de grano medio a grueso de color gris claro, en capas de 0.20 a 0.80 m de espesor, muy resistentes. La unidad cuatro presenta limolita a lutita de color gris oscuro a negro, con estratificación no definida y con ligera foliación. La unidad cinco consiste en caliza gris que intemperiza a gris claro y su espesor varía regionalmente; presenta abundantes crinoideos y gasterópodos. La unidad seis se define como una alternancia de arenisca de color café claro en capas medianas a delgadas y lutita de color gris oscuro con intercalaciones de horizontes con concreciones calcáreas. La unidad siete es la más joven y consiste en una arenisca de color negro a gris oscuro con estratificación delgada.
De acuerdo con Juárez-Arriaga (2006) las facies continentales de la Formación Olinalá están compuestas por arenisca conglomerática, conglomerado polimíctico y cuarzo-arenita que se interpretan como ambientes de depósito fluvial o costero. El mismo autor subdivide las facies marinas en carbonatadas, que incluyen biohermas de algas rojas coralináceas, packstone arcilloso de braquiópodos y wackestone/packstone de fusulínidos, packstone/grainstone de crinoideos-briozoarios-braquiópodos, entre otros; y terrígenas, compuestas por lutita de color negruzco, laminación fina, con pirita, estructuras cono-en-cono, y fósiles tales como amonoideos, nautiloideos, gasterópodos, braquiópodos, conuláridos, fragmentos desarticulados de crinoideos, restos vegetales, además de bivalvos. Por sus relaciones estratigráficas, sugieren un depósito en condiciones de plataforma externa, sin influencia de oleaje. Los ciclos en el depósito han sido considerados por el mismo autor, como el registro de tres eventos transgresivos-regresivos que ocurrieron en el área durante el Pérmico medio.
El material de bivalvos examinado se encuentra en estratos que corresponden a parte de la unidad 4 propuesta por Corona-Esquivel (1981 [1983]), y a la lutita de la secuencia SD-2 de Juárez-Arriaga (2006) (Figura 2).
Figura 2. Columna estratigráfica simplificada de la Formación Olinalá. Se indica la parte de la columna que corresponde a la unidad 4 de Corona-Esquivel (1981) y González-Arreola et al. (1994) del cual provienen los bivalvos estudiados. Modificada de Juárez-Arriaga (2006).
4. Consideraciones estratigráficas
Flores de Dios y Buitrón (1982) asignaron inicialmente a toda la Formación Olinalá una edad Pensilvánico-Triásico. Posteriormente Corona-Esquivel (1981 [1983]), Vachard et al. (1993), González-Arreola et al. (1994), Esquivel-Macías (1996) y Silva-Pineda et al. (1998, 2000) restringieron la edad al Pérmico.
Vachard et al. (2004) reconocieron una posible edad Wordiano para las capas inferiores de la Formación Olinalá basados en la presencia del amonoideo Waagenoceras sp., mientras que asignaron una edad Capitaniano a la parte carbonatada de esta formación por la presencia de los fusulínidos Polydiexodina sp. y Codonofusiella extensa; sin embargo, el alcance conocido de Polydiexodina se ha extendido, en los últimos años, del Wordiano al Capitaniano. La especie Codonofusiella extensa sólo se conoce para el Capitaniano de Coahuila, por lo que al tenerse un registro tan limitado de su distribución geográfica, tampoco puede ser considerada en este momento como un fósil índice; mientras que el género Waagenoceras presenta una distribución temporal conocida que va del Roadiano al Capitaniano.
De acuerdo a las consideraciones anteriores y a los alcances estratigráficos de los bivalvos estudiados (Figura 3), el conjunto es característico del Pérmico medio y sugiere una edad de Roadiano, lo que ampliaría la extensión del alcance estratigráfico de la especie Septimyalina burmai. Si este fuera el escenario real, habría importantes implicaciones biogeográficas, ya que en la Formación Olinalá existen especies de braquiópodos que han sido consideradas como probables formas endémicas relictas (Sour-Tovar et al., 2009).
Por otra parte, González-Arreola et al. (1994) propusieron ampliar el alcance estratigráfico de Posidoniella, un género previamente considerado del Carbonífero, hasta el Pérmico medio y posteriormente, Quiroz-Barroso et al. (2012) mencionaron la presencia de este género en rocas del Cisuraliano (Kunguriano) de la Formación Tuzancoa, en el estado de Hidalgo; con base en los resultados obtenidos en el presente estudio, se corrobora la propuesta de ampliar el alcance estratigráfico de este género hasta el Pérmico medio, por lo menos hasta el Roadiano.
Figura 3. Alcances estratigráficos de los géneros y especies estudiados. Las líneas punteadas indican el alcance estratigráfico del género; las barras negras indican el alcance estratigráfico de las especies. En el caso de Posidoniella sp., la sección con asterisco (*) indica la propuesta de González-Arreola et al. (1994) de ampliar su alcance estratigráfico hasta el Pérmico medio.
5. Consideraciones tafonómicas y paleoecológicas
Los géneros de bivalvos estudiados representan tres modos de vida principales: Nuculavus, Palaeoneilo, Edmondia y Cardiomorpha incluyen especies que fueron organismos móviles facultativos y formaron parte de la infauna; Posidoniella, Septimyalina y Leptodesma incluyen especies que vivían adheridas a diferentes sustratos formando parte de la epifauna y Bakevellia es un género con especies semi-infaunales (Figura 4). Su registro consiste en moldes internos, moldes externos y vaciados en lutita obscura, cuya preservación fue favorecida, en muchos, casos al quedar incluidos en dos tipos de concreciones.
Figura 4. Hábitos de vida de los bivalvos estudiados. Infaunales, móviles facultativos: 1, Nuculavus levatiformis (detritívoro-suspensívoro); 2, Palaeoneilo mcchesneyana (detritívoro); 3, Edmondia phosphatica (suspensívoro); 4, Cardiomorpha sp. (suspensívoro). Semi-infaunal, estacionario: 5, Bakevellia sp. (suspensívoro). Epifaunales, estacionarios: 6, Posidoniella sp. (suspensívoro); 7, Septimyalina burmai (suspensívoro); 8, Leptodesma (Leptodesma) sp. (suspensívoro). No se trata de una reconstrucción ambiental.
Tipo 1. Las que tienen concentraciones de individuos de los géneros Nuculavus y Palaeoneilo, que fueron organismos infaunales de talla pequeña y cuyo registro en aguas marinas incluye facies de costa y de plataforma (Newell et al., 1953; Sterren, 2000; Anelli et al., 2002) (Figura 5). De acuerdo a la clasificación de Kidwell et al. (1986) las concentraciones observadas en este tipo de concreciones son politípicas por la presencia de varias especies de bivalvos y braquiópodos. Su estructura bioclástica consiste en conchas pequeñas, articuladas, dispuestas en forma azarosa y sin deformación por compresión.
Figura 5. Concreciones con conchas pequeñas de braquiópodos y de los bivalvos Nuculavus levatiformis (1) y Palaeoneilo mcchesneyana (1 y 2).
Tipo 2. Aquellas en las que hay valvas desarticuladas y algunas veces incompletas de un solo individuo de los géneros Posidoniella, Septimyalina, Bakevellia o Leptodesma, que incluyen formas epifaunales o semi-infaunales que comúnmente vivían en aguas salobres; y de Edmondia o Cardiomorpha que fueron infaunales y cuyo registro en otros trabajos se asocia a depósitos de aguas poco profundas (Girty, 1910a; Ciriacks, 1963; Runnegar y Newell, 1974; Dickins, 1999).
El hecho de que los bivalvos infaunales sean abundantes en las concreciones y que presenten las valvas articuladas y, por otra parte, que los bivalvos epifaunales estén desarticulados y sus valvas muestren escaso desgaste, es congruente con los hábitos de vida de los géneros estudiados y sugiere cambios en las condiciones de depósito. Los primeros representan elementos autóctonos o parauctóctonos, mientras que los segundos que vivían a menor profundidad, como los bivalvos Posidoniella sp, especie epifaunal cuyo registro en el campo presenta valvas grandes completas, apoya la idea de un medio de baja energía y transporte moderado o rápido. Los restos vegetales asociados se interpretan como elementos alóctonos al ser transportados desde el continente hasta el mar mediante corrientes fluviales.
De acuerdo con los elementos taxonómicos y tafonómicos, y con la lutita fosilífera, se interpreta un ambiente de plataforma de aguas poco profundas de baja energía, influenciado por la desembocadura de ríos y el transporte de bivalvos provenientes de estuarios, lo cual es congruente con el modelo de Juárez-Arriaga (2006).
6. Consideraciones biogeográficas
Las reconstrucciones paleogeográficas coinciden en colocar el norte del territorio mexicano como parte de Laurusia, en una región de latitudes bajas y formando parte de un extenso mar somero (Bambach et al., 1980; Ziegler et al., 1997). Durante el Carbonífero, este mar somero tenía continuidad hacia el norte hasta la latitud más alta de América del Norte, y hacia el sur hasta Gondwana, y sólo era interrumpido por algunos terrenos poco elevados. Sin embargo, como resultado de la formación de Pangea, su extensión se redujo durante el Pérmico medio. En este mar quedó comprendida el área de estudio.
Los géneros y especies referidos presentan afinidad con faunas del Pérmico medio de América del Norte que se han relacionado con las provincias Cordillerana y Laurencia Sur del Reino Americano de acuerdo con el modelo de Bambach (1990) y con las provincias Cordillerana y Grandiana propuestas por Yancey (1975), como parte del reino Paleoecuatorial (Shen et al., 2009) (Figura 6).
Figura 6. Distribución paleogeográfica de las especies estudiadas, durante el Pérmico medio. Los números indican las especies: 1. Nuculavus levatiformis, 2. Palaeoneilo mcchesneyana, 3. Septimyalina burmai, 4. Edmondia phosphatica. Las áreas en las que se encuentra el registro de bivalvos corresponden a mares poco profundos. Reinos biogeográficos: I. Boreal; II. Paleoecuatorial y III. Gondwánico y extensión de la Provincia Grandiana (Yancey, 1975) de acuerdo a Shen et al., 2009). Mapa base modificado de Ziegler et al., 1997.
7. Paleontología sistemática
En la clasificación siguiente se adoptan la nomenclatura y clasificación propuestos por Bieler et al. (2010). Para las categorías inferiores a familia se siguen los criterios de Newell y LaRocque en Cox et al. (1969). El material descrito se encuentra depositado en la colección del Museo de Paleontología de la Facultad de Ciencias, Universidad Nacional Autónoma de México con el acrónimo FCMP.
Clase Bivalvia Linnaeus, 1758
Orden Nuculida Dall, 1889
Superfamilia Nuculoidea Gray, 1824
Familia Nuculidae Gray, 1824
Género Nuculavus Chernyshev, 1947
Especie tipo. Nuculavus minuta Chernyshev, 1947, por designación original. Carbonífero de Rusia.
Diagnosis. Nucúlidos pequeños, trigonales, con ornamentación concéntrica que consiste en líneas de crecimiento finas e irregulares, umbo ligeramente desplazado hacia la parte posterior, resilífero pequeño, abierto, poco profundo y triangular. Interior del margen ventral liso (Yancey, 1978).
Nuculavus levatiformis (Walcott, 1884)
Figuras 7.1-7.3
Nucula levatiforme Walcott, 1884, p. 241, lám. 22, fig. 1, 1a.
Nucula levatiformis Walcott. Girty in Lee y Girty 1909, p. 74, lám. 10, figs. 7, 8; Clifton, 1942, p. 693.
Palaeonucula levatiformis (Walcott) 1884. Chronic, 1952, p. 138, lám. 6, figs. 4-9; Winters, 1963, p. 49, lám. 6, figs. 4-7a; Mayou, 1967, p. 118, lám. 1, fig. 10; Yancey, 1969, p. 6, figs. 18-20.
Palaeonucula cf. levatiformis (Walcott) 1884. Chronic en Newell et al., 1953, p. 151, lám. 33, figs. 2a, b.
Nuculavus levatiformis (Walcott) 1884. Yancey, 1978, p. 316, lám. 7, figs. 6-10; Sterren, 2000, p. 428, figs. 9.A, 9.B; Anelli et al., 2002, p. 168, figs. 3c-d, 4; Sterren, 2004, p. 60, figs. 3, A-B.
Descripción. Concha pequeña, equivalva, inequilateral y subtrigonal. Dentición incompleta, con los dientes anteriores y posteriores de tamaño uniforme. Margen ventral redondeado. Carena tenue en el borde anterior. Superficie con estrías concéntricas finas y numerosas. Interior con impresiones de los músculos aductores isomiarios; también se observa la línea palial.
Material examinado. Un molde externo e interno de una valva izquierda (FCMP 841) y un molde interno de ambas valvas articuladas (FCMP 842).
Medidas. (Tabla 1)
Distribución. Nuculavus levatiformis se ha referido para el Pérmico Inferior-Medio (Asseliano-Wordiano) de Arizona, Texas, Nevada y Utah en los Estados Unidos (Clifton, 1942; Chronic, 1952; Winters, 1963; Cooper y Grant, 1972; Yancey, 1978) y en el Carbonífero Superior-Pérmico Inferior (Gzheliano–Asseliano) de Argentina (Sterren, 2000; 2004). Chronic en Newell et al. (1953) la refiere para el Paleozoico Superior (Asseliano-Artinskiano) de Perú y Anelli et al. (2002) refieren esta especie para el Pensilvánico Medio de la cuenca de Parnaíba en Brasil.
Discusión. Nuculavus se diferencía de otros géneros similares: Nuculopsis presenta valvas de forma subcuadrada y alargada (Yancey, 1978 en Anelli et al., 2002); Nucula tiene el interior del margen ventral crenulado y un resilífero que interrumpe la continuidad de la dentición (Elías, 1957 en Anelli et al., 2002).
Yancey (1978) distingue a Nuculavus levatiformis de otras especies del género Nuculavus por presentar un umbo menos prominente, valvas moderadamente convexas y márgenes anterior y posterior redondeados. El mismo autor reconoce una amplia variabilidad y diferencias regionales dentro de la especie.
Los ejemplares estudiados presentan características afines a los ejemplares mostrados por Yancey (1978) y por Anelli et al. (2002). En los ejemplares estudiados no se observa el pequeño resilífero triangular, sin embargo sí se presentan los demás caracteres de la especie.
Localidad. Camino Olinalá-El Progreso, localidad 2.
Orden Nuculanida Carter, Campbell y Campbell, 2000
Superfamilia Nuculanoidea Adams y Adams, 1858
Familia Mallettidae Adams y Adams, 1858
Género Palaeoneilo Hall y Withfield, 186
Especie tipo. Nuculites constricta Conrad, 1842, por designación subsecuente de Hall, 1885. Devónico Medio de Nueva York.
Diagnosis. Valvas posteriormente alargadas, con o sin resilífero, usualmente con seno palial. Muesca radial posterior apenas visible, ornamentación concéntrica, sin septo interno (McAlester, 1969).
Palaeoneilo mcchesneyana (Girty, 1910a)
Figuras 7.4-7.11
Yoldia mcchesneyana Girty, 1910a, p. 39, lám. 4, figs. 4-6.
Nucula mcchesneyana Branson, 1948, p. 636.
Anthraconeilo mcchesneyana Girty, 1910 a. Yancey, 1978, p. 324, lám. 8, figs. 13-15.
Palaeoneilo mcchesneyana (Girty). Ciriacks, 1963, p. 37, lám. 4, figs. 12-16; Fang, 1987, p. 364, lám. 1, figs. 8-10.
Descripción. Concha equivalva, con valvas inequilaterales, alargadas e infladas; umbo poco prominente y ortogiro desplazado hacia el centro entre los márgenes anterodorsal y posterodorsal. Margen posterodorsal más grande que el anterodorsal; margen ventral redondeado. Superficie con ornamentación comarginal consistente en líneas delgadas poco espaciadas entre sí.
Material examinado. Un molde externo de una valva derecha (FCMP 843), un molde compuesto de una valva izquierda (FCMP 844) y seis ejemplares articulados (FCMP 845-848, FCMP 849 y FCMP 850), de los cuales FCMP 849 tiene las valvas abiertas en forma de mariposa. Además se presentan varios moldes externos encontrados en el interior de concreciones de lutita Figuras 5.1 y 5.2 cuya preservación no permite medir sus dimensiones. Se trata de la segunda especie de bivalvo más abundante en las localidades estudiadas.
Medidas. (Tabla 2)
Distribución. Palaeoneilo mcchesneyana ha sido reportada para los estados de Idaho, Wyoming (Girty, 1910a; Ciriacks, 1963) y Nevada (Yancey, 1978), en Estados Unidos, y en Hunan, China (Fang, 1987). Su alcance estratigráfico abarca desde el Artinskiano hasta el Wuchiapingiano.
Discusión. Palaeoneilo mcchesneyana se diferencia de P. sera Girty (1910b) por tener el margen totalmente redondeado y no sinuoso hacia su parte posterior formando una ligera muesca, y su umbo es menos prominente (Hoare et al., 1989; Hoare, 1993); de P. tebagaensis (Termier y Termier 1959 en Boyd y Newell, 1979) por sus umbos ortogiros que no están aplanados; de P. oweni (McChesney en Hoare et al., 1979) por poseer valvas con márgenes posterodorsal y posteroventral que se juntan sin formar un contorno agudo; de P. qinzhouensis (Weihong et al., 2007) por presentar una charnela no arqueada y umbo ortogiro.
Localidad. Camino Olinalá-El Progreso, localidad 2.
Tabla 2. Dimensiones de Palaeoneilo mcchesneyana (en mm).
Orden Pteriida Newell, 1965
Superfamilia Ambonychioidea Miller, 1877
Familia Myalinidae Frech, 1891
Género Posidoniella De Koninck, 1885
Especie tipo. Inoceramus vetustus Sowerby, 1829, por designación original. Carbonífero Inferior de Inglaterra.
Diagnosis. Conchas ventricosas con el aspecto de los inocerámidos, sin ornamentación radial, con aurícula anterior. Con una delgada capa prismática (Newell en Cox et al., 1969).
Posidoniella sp.
Figuras 7.12-7.18, y Figuras 9.1, 9.2
Descripción. Valvas adultas suborbiculares, con umbo ortogiro próximo a la parte posterior que sobresale del margen dorsal. Pequeña prolongación a manera de aurícula con posición anterior. La ornamentación consiste en pliegues concéntricos gruesos y ampliamente espaciados entre sí; carecen de ornamentación radial.
Material examinado. Once moldes externos de valvas izquierdas (FCMP 851-861) y nueve de valvas derechas (FCMP 862-870), dentro de los cuales, los que presentan mejor conservación son: FCMP 851-856 y FCMP 862-864.
Medidas. (Tabla 3)
Distribución. Posidoniella ha sido referido para el Carbonífero (Viseano-Namuriano), con reportes en el Pensilvánico (Cox et al., 1969). También fue reportado para el Carbonífero Inferior de Chile por Isaacson y Dutro (1999) en el miembro superior de la Formación Las Zorritas. Para el Misisípico Superior del Reino Unido (Wolfenden, 1958) se mencionaron las especies P. gibbosa Hind 1897 y P. vetusta (Sowerby, 1829). En España se refirió para el Namuriano-Westphaliano (Sampelayo y Sampelayo, 1947). González-Arreola et al. (1994) lo refirieron para la Formación Olinalá asociado a braquiópodos (Cancrinella rugosa Cloud, 1944, Thamnosia depressa (Cooper, 1953) y Orbiculoidea ovalis Cloud, 1944), gasterópodos (Babylonites carinatus Yochelson, 1956), amonoideos (Stacheoceras toumanskyae Miller y Furnish, 1940, Paraceltites elegans Girty, 1908 y Waagenoceras dieneri (Böse, 1919), y el conulárido Paraconularia sp., que en conjunto indican una edad Pérmica. Quiroz-Barroso et al. (2012), refirieron este género para el Cisuraliano (Pérmico inferior) de Otlamalacatla, Hidalgo.
Discusión. Los ejemplares de esta especie son los más abundantes en el área, aunque su preservación no es la mejor debido a que los moldes externos se encuentran sobre láminas finas de lutita gris clara a oscura, que se rompen fácilmente al recolectar los ejemplares. El material ilustrado por González-Arreola et al. (1994) y el que se recolectó para este estudio es incompleto, pues solamente se tienen las partes próximas al umbo (Figura 8).
Debido a la pobre preservación de los ejemplares se deja abierta la determinación.
Localidad. Camino Olinalá-El Progreso, localidades 1 y 2.
Figura 8. 1, Ejemplar de Posidoniella sp. observado en el afloramiento; 2, reconstrucción del mismo ejemplar, las líneas punteadas representan las partes más frágiles de la valva que se quiebran al recolectarlas.
Tabla 3. Dimensiones de Posidoniella sp. (en mm).
Género Septimyalina Newell, 1942
Especie tipo. Myalina perattenuata Meek y Hayden, 1858, por designación original. Pensilvánico Superior de Kansas, Estados Unidos.
Diagnosis. Concha prosoclina, romboidal, falcada o aclina; delgada, con umbos extendidos y delgados, con lóbulo anterior obsoleto debajo de cada umbo; ambas valvas lisas o rugosas; inequivalvas; cavidad umbonal anterodorsalmente cubierta por una placa charnelar ancha; dentición débil, como en Myalina (Newell en Cox et al., 1969).
Septimyalina burmai (Newell, 1942)
Figuras 9.3-9.8
?Myalina apachesi Marcou, 1858, p. 44, lám. 7, figs. 6, 6a.
Myalina apachesi Marcou. Lee y Girty, 1909, p. 81, lám. 9, figs. 6, 7; Clifton, 1942, p. 690, lám. 101, figs. 17, 18; lám. 102, figs. 3, 4.
Septimyalina burmai, Newell, 1942, p. 67, lám. 12, figs. 1-6; Mudge et al., 1962, lám. 15, fig. 21; Kues, 1995, lám. 7, figs. 30-34.
Descripción. Concha mediana, mitiliforme, equivalva y prosoclina; umbo prosogiro y puntiagudo, poco prominente; margen cardinal recto, formando un ángulo obtuso con el margen posterior, que es ligeramente convexo; margen anterior convexo en la parte superior, recto en la inferior; margen ventral redondeado. Ambas valvas marcadas con laminillas concéntricas espaciadas regularmente en forma de arrugas, sin ornamentación radial.
Material examinado. Siete moldes externos de valvas derechas (FCMP 871-877), de los que el ejemplar FCMP 871 es una forma juvenil; un molde externo de una valva izquierda (FCMP 878); y dos moldes externos con las valvas articuladas (FCMP 879 y FCMP 880), de los cuales la última está abierta en forma de mariposa.
Medidas. (Tabla 4)
Distribución. Septimyalina burmai ha sido referida para el Gzheliano-Kunguriano de América del Norte. Ha sido referida para América del Norte: en Puebla, México (Sour-Tovar et al., 2000) y los estados de Kansas (Newell, 1942; Mudge et al., 1962), Nuevo México (Kues, 1995) y Oklahoma (Clifton, 1942) en Estados Unidos. En Sudamérica ha sido referido para Brasil (Anelli et al., 2006) y Perú (Newell et al., 1953).
Discusión. De acuerdo con Newell (1942), no es posible comprobar la sinonimia con el material estudiado por Marcou (1858), pues los ejemplares en que se basa el segundo trabajo se perdieron y se desconocen tanto la localidad de recolección como el horizonte exacto.
S. perattenuata (Meek y Hayden, 1858) presenta la marca del ligamento en la charnela, conformada por una región muy amplia en la concha que muestra una serie de líneas donde se inserta el ligamento. En el ejemplar estudiado, la charnela parece no ser tan grande como para que exista tal área. En el ejemplar de Newell (1942) se observa que la parte de la charnela que tiene la inserción del ligamento es muy ancha y esta área no aparece en el ejemplar estudiado. Hoare et al. (1979) incluye dentro de esta especie ejemplares con una variabilidad evidente, desde una concha muy alargada y pequeña y carente de una aurícula, hasta conchas grandes, alargadas y anchas, con una aurícula grande.
Anelli et al. (2006) presentaron varios moldes internos y un externo de S. perattenuata, de los que, en el último, se observa una notable diferencia con S. burmai: la presencia de un plegamiento en la parte antero-ventral de la valva izquierda. Dicho plegamiento no se presenta en S. burmai
Septimyalina burmai se diferencia de S. orthonota (Mather, 1915) por el umbo puntiagudo y prominente; de S. orbiculata Newell, 1942 por su charnela menos amplia, su concha menos inequilateral y su umbo menos largo; de S. sinuosa (Morningstar, 1922) por su concha menos ancha, un umbo mucho menos puntiagudo y una charnela menos amplia. S. perattenuata, S. orthonota y S. orbiculata pertenecen al Pensilvánico, y esta última pudo ser ancestro de S. scitula Newell, 1942, sin relación con S. burmai, mientras que S. burmai, S. perattenuata y S. orthonota conforman un género natural, pues de acuerdo con Newell (1942) aparecen sucesivamente en la columna geológica.
Las especies restantes del género son lo suficientemente distintas a S. burmai pues es fácil distinguir sus diferencias. Por lo tanto, con base en la comparación de los ejemplares estudiados con otras especies de este género, se determina que pertenece a Septimyalina burmai debido a que carece de aurículas, no tiene una concha ancha y sus umbos son puntiagudos y ligeramente curvados.
Localidad. Camino Olinalá-El Progreso, localidades 1 y 2.
Figura 9. 1, 2: Posidoniella sp.; 1, fragmento del margen ventral, ejemplar FCMP 864; 2, fragmentos de dos valvas, ejemplar FCMP 852; 3-8, Septimyalina burmai (Newell, 1942); 3,4, moldes externos de valvas izquierda y derecha, articuladas, abiertas en forma de mariposa, ejemplar FCMP 879; 5, molde externo de la valva izquierda, juvenil, ejemplar FCMP 871; 6, molde externo, vista anterior de ambas valvas articuladas, abiertas y deformadas, ejemplar FCMP 880; 7, molde externo de la valva izquierda, ejemplar FCMP 872; 8, molde externo de la valva izquierda, ejemplar FCMP 873; 9, Bakevellia sp. Molde externo de la valva izquierda, ejemplar FCMP 881; 10, Leptodesma (Leptodesma) sp. Molde compuesto imperfecto de la valva derecha, ejemplar FCMP 882; 11-18, Edmondia phosphatica Girty, 1910a; 11, valva izquierda, 12, valva derecha, 13, vista dorsal, 14, vista anterior, ejemplar FCMP 883; 15, valva izquierda, 16, valva derecha, 17, vista dorsal, 18, vista anterior, ejemplar FCMP 885.
Superfamilia Pterioidea Gray, 1847
Familia Bakevelliidae King, 1850
Género Bakevellia King, 1848
Especie tipo. Avicula antiqua von Münster en Goldfuss, 1836, por designación original. Pérmico Superior de Inglaterra.
Diagnosis. Concha de pequeña a mediana, de forma trapezoidal o romboidal, no muy alargada; la aurícula anterior puede ser de pequeña a moderadamente grande, el ala posterior es obtusa o muy puntiaguda; las valvas nunca son extremadamente inequivalvas; con apertura anterior estrecha. Pérmico-Cretácico, cosmopolita. (Cox et al., 1969).
Bakevellia sp.
Figura 9.9
Descripción. Valva pequeña, subcuadrada. Umbo pequeño. La aurícula anterior es angosta y puntiaguda; la aurícula posterior es amplia, de ángulo obtuso y más grande que la aurícula anterior. Ornamentación no visible.
Material examinado. Un molde imperfecto de una valva izquierda (FCMP 881).
Medidas. 3.5 mm de anchura y 2.3 mm de altura.
Distribución. El alcance estratigráfico del género Bakevellia va desde el Serpukhoviano (Pérmico) hasta el Maastrichtiano (Cretácico). Para el Pérmico, el género Bakevellia tiene reportes provenientes de Canadá (Harker y Thorsteinsson, 1960), Italia (Posenato et al., 2005), México (Blodgett et al., 2002), Reino Unido (Logan, 1967) y en Estados Unidos es donde existe una mayor cantidad de referencias provenientes de Arizona (Chronic, 1952), Idaho, Texas y Wyoming (Ciriacks, 1963), Nevada (Yancey y Stevens, 1981) y Nuevo México (Kues, 1995).
Discusión. Kues (1995) presentó numerosas conchas halladas en calizas, y las determinó como Bakevellia sulcata Geinitz (1866). En dicho trabajo se ilustran valvas izquierdas similares al ejemplar que aquí se determina; Kues (1995) indicó que se trata de valvas de tamaños menores a 5 mm. No es posible comparar la valva que aquí se determina con la de Kues, pues aunque describe a detalle sus ejemplares, sus fotografías no permiten observar bien la morfología; Winters (1963) reportó especímenes de B. sulcata cuya área ventral es mucho más larga que el ejemplar que aquí se determina, además no es posible la comparación con esta especie porque la valva que aquí se describe no presenta el sulcus, un carácter diagnóstico para la especie.
Localidad. Camino Olinalá-El Progreso, localidad 1.
Familia Pterineidae Meek, 1864
Género Leptodesma Hall, 1883
Especie tipo. Leptodesma potens Hall, 1883, por designación subsecuente de Miller, 1889. Devónico Superior de Nueva York, Estados Unidos.
Diagnosis. Sin ornamentación radial, umbo izquierdo más prominente que el derecho; ala posterior comúnmente bien desarrollada; contrafuerte miofórico débil; pocos dientes cardinales y variables en número (Newell y LaRocque en Cox et al., 1969).
Subgénero Leptodesma Hall, 1883
Leptodesma (Leptodesma) sp.
Figura 9.10
Descripción. Concha pequeña, inequivalva y poco convexa. Margen anterior ligeramente redondeado; margen posterior paralelo al anterior, convexo hacia el margen ventral; margen cardinal recto y amplio, mayor que la longitud de la concha. Umbo pequeño y poco prominente. Aurícula anterior muy reducida; aurícula posterior en forma de ala muy aguda que termina en forma de espina. Superficie ornamentada con líneas comarginales finas y espaciadas.
Material examinado. Un ejemplar (FCMP 882) que consiste en la impresión externa de la valva izquierda casi totalmente cubierta por el molde compuesto imperfecto de la valva derecha. Las valvas están desplazadas.
Medidas. 4.2 mm de anchura y 3.4 mm de altura.
Distribución. El alcance estratigráfico del subgénero Leptodesma (Leptodesma) comprende desde el Telychiano (Silúrico Inferior) al Changhsingiano (Pérmico Superior). Es cosmopolita.
Discusión. Se asigna a Leptodesma (Leptodesma) por poseer una aurícula anterior angular, que lo distingue del otro subgénero, Leptodesma (Leiopteria) Hall, 1883, en el cual es lobulada (Newell y La Rocque en Cox et al., 1969). El estado precario del único ejemplar no permite conocer características diagnósticas, por lo que se deja abierta la asignación de especie.
Localidad. Camino Olinalá-El Progreso, localidad 1.
Orden Pholadomyida Newell, 1965 [Anomalodesmata Dall, 1889]
Superfamilia Edmondioidea King, 1850
Familia Edmondiidae King, 1850
Género Edmondia De Koninck, 1841
Especie tipo. Isocardia unioniformis Phillips, 1836, por designación original. Carbonífero Inferior de Irlanda.
Diagnosis. Concha ovoide a elíptica alargada, regularmente gibosa. Sin carenas laterales o sulco; márgenes cerrados; umbos pequeños, prosogiros, situados de un cuarto a un tercio detrás del margen anterior; las valvas generalmente están ornamentadas con algunas líneas concéntricas irregulares o líneas de crecimiento; placa charnelar internamente reforzada por una cresta casi paralela (Newell en Cox et al., 1969).
Edmondia phosphatica Girty, 1910a
Figuras 9.11-9.18
Edmondia? phosphatica Girty, 1910a, p. 36, lám. 6, fig. 9.
Edmondia phosphatica Girty. Ciriacks, 1963, p. 74, lám. 11, figs. 12-15.
Descripción. Concha subcuadrada, muy inflada, inequilateral. Umbos prosogiros, muy próximos a la parte anterior. El margen anterodorsal presenta un truncamiento, mientras que el posterodorsal es largo y ocupa la mayor parte de la zona dorsal. Márgenes dorsal y ventral planos, más o menos paralelos entre sí. Charnela larga. La ornamentación es comarginal y consiste en pliegues gruesos muy pronunciados.
Material examinado. Tres moldes externos de valvas articuladas (FCMP 883, FCMP 884 y FCMP 885).
Medidas. (Tabla 5).
Distribución. Edmondia phosphatica se ha descrito para el Roadiano de Idaho y Wyoming (Girty, 1910a; Ciriacks, 1963), en Estados Unidos.
Discusión. El género Edmondia se distingue por tener una placa charnelar, que en moldes internos se observa como un surco paralelo al margen dorsal. La cara ventral de esta placa presenta un conjunto de inserciones musculares circulares, y no debe interpretarse como el sitio de fijación de un ligamento interno (Runnegar y Newell, 1974).
Edmondia phosphatica se diferencia de E. nodulifera (Hoare et al., 1978) por presentar una ornamentación con líneas gruesas; de E.? bellula Girty, 1908 por su umbo prosogiro, muy próximo a la parte anterior.
Ciriacks (1963) menciona que Edmondia phosphatica presenta valvas con ornamentación delgada; sin embargo, en la lámina 11, figuras 12-15 de dicho trabajo se observa una ornamentación más gruesa, muy similar a la que presentan los ejemplares de México. Se determina que se trata de esta especie porque los caracteres observados en el trabajo de Ciriacks coinciden con los de los ejemplares descritos aquí. También se puede observar que los ejemplares estudiados aquí tienen unos márgenes dorsal y ventral lisos y casi paralelos entre sí, lo cual concuerda con los ejemplares de Ciriacks.
Localidad. Camino Olinalá-El Progreso, localidad 2.
Superfamilia Pholadomyoidea King, 1844
Familia Grammysiidae Miller, 1877
Género Cardiomorpha De Koninck, 1844
Especie tipo. Isocardia oblonga Sowerby, 1825, por designación subsecuente de Woodward, 1854. Carbonífero Inferior de Irlanda.
Diagnosis. Similar a Edmondia, pero con umbos muy prominentes, y fuertemente prosogiros; con superficie lisa. Caracteres internos desconocidos (Newell en Cox et al., 1969).
?Cardiomorpha sp.
Figuras 10.1-10.11
Descripción. Valvas inequilaterales, ovaladas y alargadas, más largas que altas, infladas, con umbos fuertemente prosogiros, casi terminales y poco prominentes. El margen posterodorsal es mucho más amplio que el margen anterodorsal y corre casi paralelo al margen ventral; el margen ventral es ligeramente curvo, siendo mayor la curva en las partes que se aproximan al margen posteroventral y anterodorsal. Placa charnelar con una carina interna delgada y paralela al margen dorsal. No presentan lúnula, ni escudo. La ornamentación es comarginal, con líneas muy delgadas y poco espaciadas entre ellas.
Material examinado. Once moldes externos con las valvas articuladas y cerradas (FCMP 886-896). De éstos, los ejemplares FCMP 895 y FCMP 896 presentan un desgaste que impide medir sus dimensiones.
Medidas. (Tabla 6).
Distribución. El registro del género Cardiomorpha abarca desde el Emsiano Tardío (Devónico) hasta el Wuchiapingiano (Pérmico Tardío). En el Reino Unido ha sido reportado para el Asbiano (Moore, 1958) y Brigantiano (Wolfenden, 1958); en China se refirió para el Eifeliano (Zhang y Pojeta, 1986) y el Wuchiapingiano (Zhao y Cheng, 1988); en India se reportó para el Sakmariano (Reed, 1932); en la Antártida se ha referido para el Serpukhoviano-Bashkiriano (Kelly et al., 2001); en Brasil se reportó para el Emsiano (Melo, 1988) y el Eifeliano-Frasniano (Morsch, 1986); en Estados Unidos se refirió para el Serpukhoviano de Virginia del Oeste (Hoare, 1993), el Desmoinesiano de Missouri (Hoare, 1961), Pensilvánico de Utah (Chamberlain, 1981), Misisípico de Alaska (Buddington y Chapin, 1929), Roadiano de Idaho (Girty, 1910a) y el Merameciano de Nevada (Brew y Gordon, 1971) y de Arkansas (Branson, 1958).
Discusión. El material examinado presenta similitud con los géneros Edmondia y Cardiomorpha debido a que ambos géneros tienen valvas ovaladas o subcuadradas y umbos prosogiros. Sin embargo, se diferencian en que los umbos de Edmondia están más desplazados hacia el centro ubicándose a un tercio desde el margen anterodorsal hacia el margen posterodorsal, mientras que los umbos de Cardiomorpha son casi terminales. Se determinó que los ejemplares revisados pertenecen al género Cardiomorpha por presentar éste último carácter.
El material examinado presenta afinidad con el material estudiado por Girty (1910a), quien describió un ejemplar de Cardiomorpha sp. como una concha con forma transversalmente subelíptica, con un umbo no muy prominente y casi terminal, con extremos simétricamente redondeados, y una superficie marcada por muchas estrías tenues y finas.
Pojeta (1969), al revisar los bivalvos de Girty (1910b), los asignó con duda a la especie Cardiomorpha? inflata, cuyos caracteres son distintivos del género, pero se distingue del material examinado principalmente en que su margen posterior es subrectilíneo y fuertemente oblicuo, haciendo que las valvas se observen alargadas y oblicuas, carácter del que carecen los ejemplares examinados, pues éstos tienen el margen posterior redondeado y, aunque sus valvas están alargadas, no son oblicuas.
La especie Cardiomorpha missouriensis presentada en Hoare et al. (1979) tiene el margen posterior subtruncado, mientras que el margen posterior del material revisado es redondeado, por lo tanto, no son equiparables. En ese trabajo se muestra la charnela del ejemplar que allí se determina; ésta tiene unas ninfas bien desarrolladas y una placa charnelar que presenta una delgada cresta interna que corre paralela a ella. En el material que se revisó hay unos ejemplares en los que las valvas están desplazadas; allí se expone una parte de la charnela cuya forma es similar a la que aparece en Hoare et al. (1979), aunque no se observa la cresta interna, lo cual se puede explicar por el hecho de que se rellenó de sedimento durante su fosilización y se perdió el carácter.
Al comparar el material revisado con ejemplares de Cardiomorpha obovata presentados en Hoare (1993), se observan grandes diferencias, pues los ejemplares de Hoare tienen los umbos muy desplazados hacia el centro, y el mismo autor los describe como umbos prosogiros, localizados aproximadamente a un cuarto de longitud desde el margen anterior; al observar las figuras, los ejemplares tienen más afinidad con formas parecidas a Edmondia. En el mismo trabajo aparecen dos ejemplares determinados como Cardiomorpha sp., los cuales sí presentan características afines al género, pero son distintos al material revisado porque sus valvas son más alargadas y con una forma más subcuadrada, además tienen un margen anterior liso y convexo hacia su margen anteroventral, caracteres que no presenta el material examinado.
Localidad. Camino Olinalá-El Progreso, localidad 2.
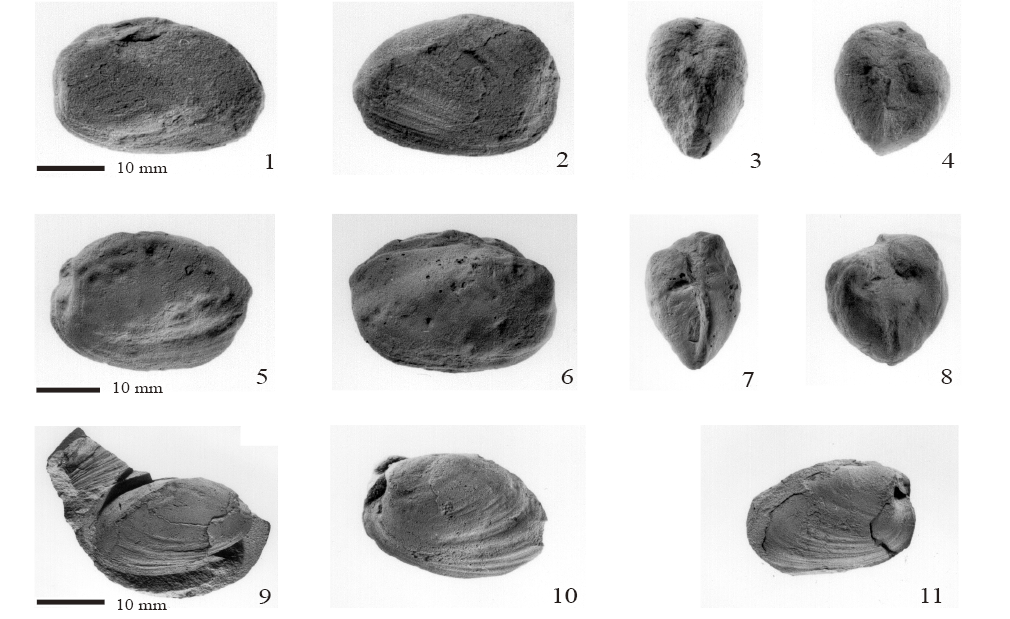
Figura 10. 1-11: ?Cardiomorpha sp.; 1, valva izquierda, 2, valva derecha, 3, vista dorsal, 4, vista anterior, ejemplar FCMP 888; 5, valva izquierda, 6, valva derecha, 7, vista dorsal, 8, vista anterior, ejemplar FCMP 894; 9, valva derecha, ejemplar FCMP 896; 10, valva izquierda, ejemplar FCMP 892; 11, valva derecha, ejemplar FCMP 893.
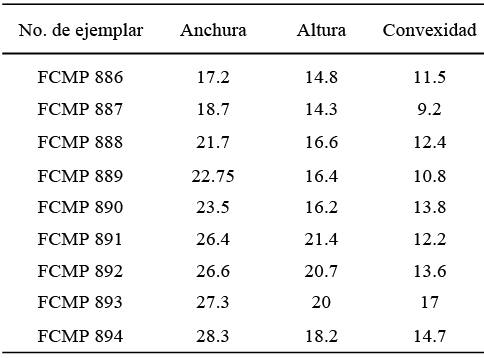
8. Conclusiones
Los bivalvos estudiados provienen de la lutita media de la Formación Olinalá, que corresponde a la unidad 4 de Corona-Esquivel (1981 [1983]) y González-Arreola et al. (1994), y a la parte lutítica de la secuencia SD-2 de Juárez-Arriaga (2006).
Se identificaron cuatro especies: Nuculavus levatiformis, Palaeoneilo mcchesneyana, Septimyalina burmai y Edmondia phosphatica. Tres especies sólo pudieron ser determinadas a nivel de género: Posidoniella sp., Bakevellia sp. y Leptodesma (Leptodesma) sp. Una especie se relacionó con duda al género Cardiomorpha.
Las especies estudiadas confirman la edad Pérmico medio (Guadalupiano) de la Formación Olinalá y sugieren una posible edad Roadiano para el nivel estudiado.
Los géneros encontrados representan tres modos de vida principales: Nuculavus, Palaeoneilo, Edmondia y Cardiomorpha incluyen especies que fueron organismos móviles facultativos y formaron parte de la infauna; Posidoniella, Septimyalina y Leptodesma incluyen especies que vivían adheridas a diferentes sustratos formando parte de la epifauna y Bakevellia es un género con especies semi-infaunales. Los cuatro últimos géneros han sido asociados comúnmente a aguas costeras y salobres. Estas diferencias ecológicas y las características particulares de las concentraciones fósiles y de la roca portadora, indican que la lutita donde se hallaron los fósiles corresponde a un ambiente de plataforma de aguas poco profundas de baja energía, influenciado por la desembocadura de ríos.
La afinidad que tienen las especies de bivalvos estudiadas con las faunas de la parte central y centro-oeste de Norteamérica, indica una relación con las provincias Cordillerana y Laurencia Sur como parte del Reino Americano de Bambach (1990). De acuerdo a las provincias pérmicas de Yancey (1975) corresponden a las provincias Grandiana y Cordillerana, consideradas por Shen et al. (2009) dentro del Reino Paleoecuatorial. Esta afinidad representa la continuidad del mar somero que cubrió la región de Olinalá con el mar que se extendió en la parte centro-oeste del norte de América durante el Pérmico medio.
Agradecimientos
Los autores agradecen a la Universidad Nacional Autónoma de México su apoyo a esta investigación a través de los proyectos PAPIIT IN 213710 e IN215013; a Leonora Martín-Medrano por la elaboración de los moldes en silicón; a Daniel Navarro-Santillán, Edgar Juárez-Arriaga y Francisco Sour-Tovar por su gran ayuda en el trabajo de campo y a Héctor Hernández-Campos por la elaboración de las fotografías del material estudiado. También, se agradece a María del Carmen Perrilliat (Instituto de Geología, UNAM) y a Luiz Eduardo Anelli (Universidade de Sao Paulo, Brasil) su cuidadosa revisión y críticas constructivas para mejorar el manuscrito.
Referencias
Adams, H., Adams, A., 1858, The genera of recent Mollusca: arranged according to their organization: London, John Van Voorst, 3, 484 p.
Anelli, L.E., Rocha-Campos, A.C., Simões, M.G., 2002, Protobranch bivalves from the Piauí Formation (Middle Pennsylvanian), Parnaíba Basin, Brazil: Revista Española de Paleontología, 17, 165-176.
Anelli, L.E., Rocha-Campos, A.C., Simões, M.G., 2006, Pennsylvanian pteriomorphian bivalves from the Piauí Formation, Parnaíba basin, Brazil: Journal of Paleontology, 80(6), 1125-1141.
Bambach, R.K., 1990, Late Palaeozoic provinciality in the Marine Realm, en McKerrow, W.S., Scotese, C.R., (eds.), Palaeozoic Palaeogeography and Biogeography: Geological Society Memoir, 12, 307-323.
Bambach, R.K., Scotese, C.R., Ziegler, A.M., 1980, Before Pangea: The Geographies of the Paleozoic World: American Scientist, 68, 26-38.
Bieler, R., Carter, J.G., Coan, E.V., 2010, part. 2, Classification of Bivalve Families, en Bouchet, P., Rocroi, .J.-P., Bieler, R., Carter, J.G., Coan, E.V. (eds.), Nomenclator of bivalve families with a Classification of Bivalve Families: Malacologia, 52(2), 1-184.
Blodgett, R.B., Moore, T.E., Gray, F., 2002, Stratigraphy and Paleontology of Permian Lower Rocks of north Cananea, northern Sonora, Mexico: Journal of South American Earth Sciences, 15, 481-495.
Böse, E., 1919, The Permo-Carboniferous ammonoids of the Glass Mountains, west Texas, and their stratigraphic significance: University of Texas Bulletin, 1762, 1-241.
Boyd, D.W., Newell, N.D., 1979, Permian pelecypods of Tunisia: American Museum Novitates, 2686, 1-22.
Branson, C.C., 1948, Bibliographic index of Permian invertebrates: Geological Society of America Memoir, 26, 1-1049.
Branson, C.C., 1958, Two Mississippian species of Conocardium: Oklahoma Geology Notes, 18, 137-142.
Brew, D.A., Gordon, M., 1971, Mississippian stratigraphy of the Diamond Peak Area, Eureka County, Nevada: United States Geological Survey Professional Paper, 661, 1-84.
Buddington, A.F., Chapin, T., 1929, Geology and mineral deposits of southeastern Alaska: United States Geological Survey Bulletin, 800, 1-398.
Carter, J.G., Campbell, D.C., Campbell, M.R., 2000, Cladistic perspectives on early bivalve evolution, en Harper, E.M., Taylor, J.D., Crame, J.A. (eds.), The evolutionary biology of the Bivalvia: Geological Society Special Publication, 177, 1-494.
Chamberlain, A.K., 1981, Biostratigraphy of the Great Blue Formation: Bringham Young University Geology Studies, 28(3), 9-17.
Chernyshev, B.I., 1947, Dekayi Nuculidae Donets´kogo basenyu: Akademie Nauk Ukrains´koyi RSR (Kiev), Instytut Geologichnykh Nauk, Zbirnyk prats´z paleontologiyi ta Stratigrafiyi, 1, 3-19.
Chronic, H., 1952, Molluscan fauna from the Permian Kaibab Formation, Walnut Canyon, Arizona: Geological Society of America Bulletin, 63(2), 95-166.
Ciriacks, K.W., 1963, Permian and Eotriassic bivalves of the Middle Rockies: Bulletin of the American Museum of Natural History, 125(1), 1-100.
Clifton, R.L., 1942, Invertebrate faunas from the Blaine and the Dog Creek Formations of the Permian Leonard Series: Journal of Paleontology, 16(6), 685-699.
Cloud, P.E., 1944, Geology and paleontology of the Permian area northwest of Las Delicias, southwestern Coahuila, Mexico, Permian brachiopods: Geological Society of America Special Paper, 52, 49-69.
Conrad, T.A., 1842, Observations on the Silurian and Devonian systems of the United States, with descriptions of new organic remains: Journal of the Academy of Natural Sciences of Philadelphia, 8, 228-280.
Cooper, G.A., 1953, Sponges, Brachiopoda, Pelecypoda, and Scaphopoda. Permian fauna at El Antimonio, western Sonora, Mexico: Smithsonian Miscellaneous Collections, 119(2), 21-80.
Cooper, G.A., Grant, R.E., 1972, Permian brachiopods of west Texas, I: Smithsonian Contributions to Paleobiology, 14, 231 p.
Corona-Esquivel, R., 1981 (1983), Estratigrafía de la región de Olinalá-Tecocoyunca, noreste del estado de Guerrero: Revista del Instituto de Geología, 5(1), 17-24.
Corona-Esquivel, R., 1985, Geología de la región comprendida entre Olinalá y Huamuxtitlán, Estado de Guerrero: Ciudad de México, Universidad Nacional Autónoma de México, tesis de licenciatura, 99 p.
Cox, L.R., Newell, N.D., Boyd, D.W., Branson, C.C, Casey, R., Chavan, A., Coogan, A.H., Dechaseaux, C., Fleming, C.A., Haas, F., Hertlein, L.G., Kauffman, E.G., Keen, A.M., LaRocque, A., McAlester, A.L., Moore, R.C., Nuttall, C.P., Perkins, B.F., Puri, H.S., Smith, L.A., Sooth-Ryen, T., Stenzel, H.B., Trueman, E.R., Turner, R.D., Weir, J., 1969, Treatise on Invertebrate Paleontology, part N Mollusca, 6. Bivalvia: Lawrence, The Geological Society of America and The University of Kansas Press, 952 p.
Dall, W.H., 1889, On the hinge of pelecypodes and its development, with an attempt toward a better subdivision of the group: American Journal of Science, 38, 445-462.
Dickins, J.M., 1999, Mid-Permian (Kubergandian-Murgabian) bivalves from the Khuff Formation, Oman: implications for world events and correlation: Rivista Italiana di Paleontologia e Stratigrafia, 105(1), 23-36.
Erben, H.K., 1956, El Jurásico Inferior de México y sus amonitas: XX Congreso Geológico Internacional, México, Universidad Nacional Autónoma de México, Instituto de Geología, 393 p.
Esquivel-Macías, C., 1996, Invertebrados del Paleozoico Tardío de las regiones de Olinalá, Gro. y Patlanoaya, Pue.: Ciudad de México, Universidad Nacional Autónoma de México, tesis de maestría, 82 p.
Esquivel-Macías, C., 2000, Braquiópodos y crinoides del Paleozoico Tardío de las Formaciones Olinalá, Gro.; Patlanoaya y Cuxtepeque, Pue.: Ciudad de México, Universidad Nacional Autónoma de México, tesis de doctorado, 124 p.
Esquivel-Macías, C., Solís-Marín, F., Buitrón-Sánchez, B.E., 2004, Nuevos registros de placas columnares de crinoideos (Echinodermata, Crinoidea) del Paleozoico Superior de México, algunas implicaciones paleobiogeográficas y paleoambientales: Coloquios de Paleontología, 54, 15-23.
Fang, Z.J., 1987, Bivalves from the Upper part of the Permian in southern Hunan, China: Collection of Postgraduate Theses of the Nanjing Institute of Geology and Palaeontology, Academia Sinica, 1, 349-411.
Flores de Dios, A., 1986, Estratigrafía de la Montaña de Guerrero entre los límites con Puebla y Oaxaca: Ciudad de México, Instituto Politécnico Nacional, tesis de licenciatura, 85 p.
Flores de Dios, A., Buitrón, B.E., 1982, Revisión y aportes a la estratigrafía de la Montaña de Guerrero: Universidad Autónoma de Guerrero, Serie Técnico-Científica, 12, 28 p.
Flores de Dios, A., Vachard, D., Buitrón, B.E., 2000, La cubierta sedimentaria pérmica superior (formaciones Olinalá, Ihualtepec y Yododeñe) de los terrenos Zapoteco y Mixteco, evolución sedimentológica e interpretaciones paleogeográficas (resumen), en II Reunión Nacional de Ciencias de la Tierra, Puerto Vallarta, Jalisco, México: Unión Geofísica Mexicana, GEOS, Resúmenes y Programa, 20(3), 21-22.
Frech, F., 1891, Die devonischen Aviculiden Deutschlands, ein Beitrag zur Systematik und Stammesgeschichte der Zweischaler: Abhandlungen zur geologischen Specialkarte von Preussen und Thüringischen Staaten, 9, 253 p.
García-Díaz, J.L., Marc, T., Campa-Uranga, M.F., Henrriette, L., 2000, Las Ignimbritas de Olinalá, Guerrero (Sierra Madre del Sur): Testigos de un margen activo continental Pacífico durante el Triásico?-Jurásico Medio, en II Reunión Nacional de Ciencias de la Tierra, Puerto Vallarta, Jal., Unión Geofísica Mexicana, GEOS, Resúmenes y Programa, 20(3), 329-330.
Garibay-Romero, L.M., 2000, Nuevos amonitas pérmicos de la Formación Olinalá-Los Arcos, Olinalá, Gro., México (resumen), en II Reunión Nacional de Ciencias de la Tierra, Puerto Vallarta, Jal., Unión Geofísica Mexicana, GEOS, Resúmenes y programa, 20(3), 299.
Geinitz, H.B., 1866, Carbonformation und Dyas in Nebraska: Kaiserliche deutsche Akademie Naturforscher zu Halle: Nova Acta Leopoldina, Dresden, Verhandlung, 33, 91 p.
Girty, G.H., 1908, The Guadalupian fauna: United States Geological Survey Professional Paper, 58, 1-651.
Girty, G.H., 1910a, The fauna of the phosphate beds of the Park City Formation in Idaho, Wyoming and Utah: United States Geological Survey Bulletin, 436, 82 p.
Girty, G.H, 1910b, New genera and species of Carboniferous fossils from the Fayetteville shale of Arkansas: Annals of the New York Academy of Sciences, 20(3) part 2, 189-238.
Goldfuss, G.A., 1833-1840, Petrefacta Germaniae: Arus and Company, 312 p.
González-Arreola, C., Villaseñor-Martínez, A.B., Corona-Esquivel, R., 1994, Permian fauna of the Los Arcos Formation, Municipality of Olinalá, State of Guerrero, Mexico: Revista Mexicana de Ciencias Geológicas, 11(2), 214-221.
Gray, J.E., 1824, Conchological observations, being an attempt to fix the study of conchology on a firm basis: The Zoological Journal, 1(2), 204-223.
Gray, J.E., 1847, A list of the genera of Recent Mollusca, their synonyms and types: Proceedings of the Zoological Society of London, 15, 129-219.
Hall, J., 1883, Lamellibranchiata, plates and explanations: New York Geological Survey Paleontology, 5(1), 20 p.
Hall, J., 1885, Lamellibranchiata II, descriptions and figures of the Dimyaria of the Upper Heidelberg, Hamilton, Portage and Chemung groups: New York Geological Survey Paleontology, 5, pt. 1, 269-562.
Hall, J., Whitfield, R.P., 1869, Preliminary notice of the lamellibranchiate shells of the upper Heidelberg, Hamilton and Chemung groups, with others from the Waverly sandstones, pt. 2, 80 p.
Harker, P., Thorsteinsson, R., 1960, Permian rocks and faunas of Grinnell Peninsula, Arctic Archipelago: Geological Survey of Canada Memoir, 309, 89 p.
Hind, W., 1896-1900, A monograph of the British Carboniferous Lamellibranchiata, 1. (1898): 1 (3): Nuculidae, Trigonidae, Unionidae, Edmondidae: Monograph of the Palaeontographical Society, 52, 209-276.
Hoare, R.D., 1961, Desmoinesian Brachiopoda and Mollusca from southwest Missouri: Missouri University Studies, 36, 262 p.
Hoare, R.D., 1993, Mississippian (Chesterian) bivalves from the Pennsylvanian Stratotype area in West Virginia and Virginia: Journal of Paleontology, 67(3), 374-396.
Hoare, R.D., Sturgeon, M.T., Kindt, E.A., 1978, New Pennsylvanian bivalves and occurrences of Pseudoconocardium from Ohio and West Virginia: Journal of Paleontology, 52(5), 1023-1036.
Hoare, R.D., Sturgeon, M.T., Kindt, E.A., 1979, Pennsylvanian marine Bivalvia and Rostroconchia of Ohio: Ohio Geological Survey Bulletin, 67, 77 p.
Hoare, R.D., Heaney III, M.J., Mapes, R.H., 1989, Bivalves (Mollusca) from the Imo Formation (Mississippian, Chesterian) of North-Central Arkansas: Journal of Paleontology, 63(5), 582-603.
Isaacson, P., Dutro, T., 1999, Lower Carboniferous brachiopods from Sierra de Almeida, northern Chile: Journal of Paleontology, 73(4), 625-633.
Juárez-Arriaga, E., 2006, Marco de Estratigrafía de Secuencias para la Sucesión Sedimentaria Terrígeno-Carbonatada del Paleozoico Superior del área de Olinalá, Guerrero, México: México, D.F., Universidad Nacional Autónoma de México, tesis de maestría, 93 p.
Kelly, S.R., Doubleday, P.A., Brunton, C.H., Dickins, J.M., Sevastopulo, G.D., Taylor, P.D., 2001, First Carboniferous and Permian marine macrofaunas from Antarctica and their tectonic implications: Journal of the Geological Society, 158(2), 219-232.
Kidwell, S.M., Fürsich, F.T., Aigner, T., 1986, Conceptual Framework for the Analysis and Classification of Fossil Concentrations: Palaios, 1, 228-238.
King, W., 1844, On a new genus of Palaeozoic shells: Annals and Magazine of Natural History, 14(92), 313-317.
King, W., 1848, A catalogue of the organic remains of the Permian rocks of Northumberland and Durham, Newcastle upon Tyne.
King, W., 1850, A monograph of the Permian fossils of England: Palaeontographical Society, XXXVIII, 258 p.
Koninck, L.G., De, 1841-1844, Description des animaux fossiles qui se trouvent dans le terrain carbonifère de Belgique. H. Dessain, Liège, 650 p.
Koninck, L.G., De, 1885, Marine Fauna of the Early Permian (Wolfcampian) Robledo Mountains Member, Hueco Formation, Southern Robledo Mountains, New Mexico, en Lucas, S.G., Heckert, A.B. (eds.), Early Permian footprints and facies: New Mexico Museum of Natural History and Science Bulletin, 6, 63-90.
Kues, B.S., 1995, Marine Fauna of the Early Permian (Wolfcampian) Robledo Mountains Member, Hueco Formation, Southern Robledo Mountains, New Mexico, en Lucas, S.G., Heckert, A.B. (eds.), Early Permian footprints and facies: New Mexico Museum of Natural History and Science Bulletin, 6, 63-90.
Lee, T., Girty, G.H., 1909, The Manzano Group of the Rio Grande Valley: United States Geological Survey Bulletin, 389, 120 p.
Linnaeus, C., 1758, Systema Naturae per regna tria naturae. Secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis, Tomus I.: Stockholm, Laurentii Salvii, Editio decima, reformata, 824 p.
Logan, A., 1967, The Permian bivalvia of northern England: Palaeontographical Society Monographs, 121, 72 p.
Marcou, J., 1858, Geology of North America with two reports on the Prairies of Arkansas and Texas, the Rocky Mountains of New Mexico, and the Sierra Nevada of California: Zürcher and Furrer, 144 p.
Mather, K.F., 1915, The Fauna of the Morrow Group of Arkansas and Oklahoma: Denison University Scientific Laboratories Bulletin, 18, 59-284.
Mayou, T.V., 1967, Paleontology of the Permian Loray Formation in White Pine County, Nevada: Brigham Young University Research Studies, Geology Series, 14, 101-122.
McAlester, A.L., 1969, Family Praenuculidae, en Cox, L.R., Newell, N.D., Branson, C.C., Casey, R., Chavan, A., Coogan, A.H., Dechaseaux, C., Fleming, C.A., Haas, F., Hertlein, L.G., Keen, A.M., LaRocque, A., McAlester, A.L., Perkins, B.F., Puri, H.S., Smith, L.A., Soot-Ryen, T., Stenzel, H.B., Turner, R.D., Weir, J., Treatise on Invertebrate Paleontology, Part N, Mollusca 6 (Bivalvia), N229, University of Kansas Press and Geological Society of America, United States of America.
Meek, F.B., 1864, Remarks on the family Pteriidae (=Aviculidae) with descriptions of some new fossil genera: The American Journal of Science and Arts, ser. 2, 37, 212-220.
Meek, F.B., Hayden, F.V., 1858, Descriptions of new organic remains collected in Nebraska territory together with some remarks on the geology of the Black Hills and portions of the surrounding country: Proceedings of the Academy of Natural Sciences of Philadelphia, 8, 70-72.
Melo, J.H., 1988, The Malvinokaffric realm in the Devonian of Brazil, en McMillan, N.J., Embry, A.F., Glass, D.J., (eds.), Devonian of the World: Canadian Society of Petroleum Geologists, Calgary, Alberta, Canada, I, 669-703.
Miller, S.A., 1877, The American Palaeozoic fossils: (publicado por el autor), Cincinnati, Ohio, xv + 253 p.
Miller, S.A., 1889, North American geology and palaeontology, 664 p.
Miller, A.K., Furnish., W.M., 1940, Permian ammonoids of the Guadalupe Mountain region and adjacent area: Geological Society of America Special Paper, 26, 1-242.
Moore, D., 1958, The Yoredale Series of Upper Wensleydale and adjacent parts of North-West Yorkshire: Proceedings of the Yorkshire Geological Society, 31(2), 91-148.
Morningstar, H., 1922, Pottsville fauna of Ohio: Ohio Geological Survey Bulletin, 25, 312 p.
Morsch, S.M., 1986, Bivalves (Mollusca) na Formação Ponta Grossa (Bacia do Paraná-Devoniano): Revisão Sistemática: Anais da Academia Brasileira de Ciências, 58(3), 403-431.
Mudge, M.R., Yochelson, E.L., Douglas, R.C., Duncan, H., Strimple, H.L., Gordon, M., Dunkler, D.H., 1962, Stratigraphy and Paleontology of Uppermost Pennsylvanian and Lowermost Permian rocks in Kansas: United States Geological Survey Professional Paper, 323, 209 p.
Newell, N.D., 1942, Late Paleozoic Pelecypods: Mytilacea: University of Kansas Publications, 10(2), 80 p.
Newell, N.D., 1965, Classification of the Bivalvia: American Museum Novitates, 2206, 25 p.
Newell, N.D., Chronic, J., Roberts, T.G., 1953, Upper Paleozoic of Peru: The Geological Society of America Memoir, 58, 276 p.
Ortega-Gutiérrez, F., Elías-Herrera, M., Reyes-Salas, M., Macías-Romo, C., López, R., 1999, Late Ordovician-Early Silurian continental collisional orogeny in southern Mexico and its bearing on Gondwana-Laurentia connections: Geology, 27, 719-722.
Phillips, J., 1836, Illustrations of the geology of Yorkshire, part. 2, the Mountain Limestone District, London, John Murray, 253 p.
Pojeta Jr., J., 1969, Revision of some of Girty’s invertebrate fossils from the Fayetteville shale (Mississippian) of Arkansas and Oklahoma-Pelecypods: United States Geological Survey Professional Paper, 606-C, 15-24.
Posenato, R., Pelikán, P., Hips, K., 2005, Bivalves and Brachiopods near the Permian-Triassic boundary from the Bükk Mountains (Bálvány-North Section, northern Hungary): Rivista Italiana di Paleontología e Stratigrafia, 111(2), 215-232.
Quiroz-Barroso, S.A., Sour-Tovar, F., Centeno-García, E., 2012, Gasterópodos y bivalvos cisularianos (Pérmico inferior) de Otlamalacatla, Hidalgo, México: Revista Mexicana de Ciencias Geológicas, 29(1), 158-178.
Reed, F.R., 1932, New fossils from the agglomerate Slate of Kashmir: Palaeontological Indica, Memoirs of the Geological Survey of India, 20(1), 79 p.
Runnegar, B., Newell, N.D., 1974, Edmondia and the Edmondiacea Shallow-Burrowing Paleozoic Pelecypods: American Museum Novitates, 2533, 19 p.
Sampelayo, P., Sampelayo, A., 1947, Fauna Carbonífera de Villablino (León): Notas y Comunicaciones del Instituto Geológico y Minero de España, 17, 3-24.
Servicio Geológico Mexicano (SGM), 2006, Léxico Estratigráfico de México http://www.sgm.gob.mx, consultado el 10 de octubre de 2012.
Shen, S.Z., Xie, J.F., Zhang, H., Shi, G.R., 2009, Roadian–Wordian (Guadalupian, Middle Permian) global palaeobiogeography of brachiopods: Global and Planetary Change, 65(3-4), 166-181.
Silva-Pineda, A., Buitrón, B.E., Flores de Dios, A., 1998, Biota (continental y marina) del Pérmico de la región de Olinalá, estado de Guerrero (resumen), en 1ª Reunión Nacional de Ciencias de la Tierra, Puerto Vallarta, Jalisco, México: GEOS, Unión Geofísica Mexicana, Resúmenes y Programa, 55 p.
Silva-Pineda, A., Buitrón, B.E., Flores de Dios, A., 2000, Presencia de coníferas en el Pérmico (Leonardiano) de Calnalli, Hidalgo; Patlanoaya, Puebla; y Olinalá, Guerrero (resumen), en 2ª Reunión Nacional de Ciencias de la Tierra, Puerto Vallarta, Jalisco, México: GEOS, Unión Geofísica Mexicana, Resúmenes y Programa, 20(3), 298-299.
Sour-Tovar, F., Quiroz-Barroso, S.A., Applegate, S.P., 2000, Presence of Helicoprion (Chondrichthyes, Elasmobranchii) in the Permian Patlanoaya Formation, Puebla, Mexico: Journal of Paleontology, 74(2), 363-366.
Sour-Tovar, F., Juárez-Arriaga, E., Murillo-Muñetón, G., 2009, Nuevos braquiópodos para el Pérmico del área de Olinalá, Guerrero, México: XI Congreso Nacional de Paleontología, 73.
Sowerby, J., 1823-1825, The mineral conchology of Great Britain; or coloured figures and descriptions of those remains of testaceous animals or shells, which have been preserved at various times and depths in the earth: London, Vol. V, 168 p.
Sowerby, J., 1829, The mineral conchology of Great Britain; or coloured figures and descriptions of those remains of testaceous animals or shells, which have been preserved at various times and depths in the earth: London, Vol. VI, 250 p.
Sterren, A.F., 2000, Moluscos bivalvos en la Formación Río del Peñón, Carbonífero tardío-Pérmico temprano, provincia de La Rioja: Ameghiniana, 37(4), 421-438.
Sterren, A.F., 2004, Bivalvos pérmicos de la Formación Tupe en la quebrada de La Herradura, provincia de San Juan: Ameghiniana, 41(1), 57-74.
Vachard, D., Oviedo, A., Flores de Dios, A., Malpica, R., Brunner, P., Guerrero, M., Buitrón, B.E., 1993, Barranca d’Olinalá (Guerrero): Une coupe de réference pour le Permien du Mexique central; étude préliminaire: Annales de la Société Géologique du Nord, 2, 155-162.
Vachard, D., Flores de Dios, A., Buitrón, B.E., 2004, Guadalupian and Lopingian (Middle and Late Permian) deposits from Mexico and Guatemala, a review with new data: Geobios, 37, 99-115.
Walcott, Ch.D., 1884, Paleontology of the Eureka District: United States Geological Survey Monograph, 8, 298 p.
Weihong, H., Feng, Q., Weldon, E.A., Gu, S., Meng, Y., Zhang, F., Wu, S., 2007, A Late Permian to Early Triassic Bivalve fauna from the Dongpan Section, Southern Guangxi, South China: Journal of Paleontology, 81(5), 1009-1019.
Winters, S.S., 1963, Supaí Formation (Permian) of eastern Arizona: Geological Society of America Memoir, 89, 99 p.
Wolfenden, E.B., 1958, Paleoecology of the Carboniferous reef complex and shelf limestones in the northwest Derbyshire, England: Bulletin of the Geological Society of America, 69, 871-898.
Woodward, S.P., 1854, A manual of the Mollusca being a treatise on recent and fossil shells: London, Crosby Lockwood and Co., 518 p.
Yancey, T.E., 1969, A molluscan faunal assemblage from the Arcturus Formation, Nevada: PaleoBios, 8, 20 p.
Yancey, T.E., 1975, Permian marine biotic provinces in North America: Journal of Paleontology, 49(4), 758-766.
Yancey, T.E., 1978, Brachiopods and Molluscs of the Lower Permian Arcturus Group, Nevada and Utah, part 1: brachiopods, scaphopods, rostroconchs, and bivalves: Bulletins of American Paleontology, 74(303), 257-367.
Yancey, T.E., Stevens, C.H., 1981, Early Permian fossil communities in northeastern Nevada and northwestern Utah, en Gray, J., Boucot, A.J., Berry, W.B.N., (eds.), Communities of the Past: Hutchinson Ross Publishing Co., Stroudsburg, Pennsylvania, 243-268.
Yochelson, E.L., 1956, Permian Gastropoda of the southwestern United States. 1. Euomphalacea, Trochonematacea, Pesudophoracea, Anomphalacea, Craspedostomatacea, and Platyceratacea: Bulletin of the American Museum of Natural History, 110(3):173-276.
Zhang, R., Pojeta, J., 1986, Devonian sections in Guangxi from which pelecypods were collected. Devonian rocks and Lower and Middle pelecypods of Guangxi, China and the Traverse Group of Michigan: United States Geological Survey Professional Paper, 1394-E, 33-43.
Zhao, J.M., Cheng, Z.X., 1988, A tabulate coral from the Lower Permian Toeling Formation at Douling of Changning County, Hunan, China: Acta Palaeontologica Sinica, 27, 650-652.
Ziegler, A.M., Hulver, M.L., Rowley, D.B., 1997, Permian world topography and climate, en Martini, I.P., (ed.), Late glacial and postglacial environmental changes: Quaternary, Carboniferous-Permian and Proterozoic: Oxford University, 111-146.
Manuscrito recibido: Junio 4, 2013.
Manuscrito corregido recibido: Agosto 4, 2013.
Manuscrito aceptado: Agosto 7, 2013.
Superfamilia Pholadomyoidea King, 1844
Familia Grammysiidae Miller, 1877
Género Cardiomorpha De Koninck, 1844
Especie tipo. Isocardia oblonga Sowerby, 1825, por designación subsecuente de Woodward, 1854. Carbonífero Inferior de Irlanda.
Diagnosis. Similar a Edmondia, pero con umbos muy prominentes, y fuertemente prosogiros; con superficie lisa. Caracteres internos desconocidos (Newell en Cox et al., 1969).
?Cardiomorpha sp.
Figuras 10.1-10.11
Descripción. Valvas inequilaterales, ovaladas y alargadas, más largas que altas, infladas, con umbos fuertemente prosogiros, casi terminales y poco prominentes. El margen posterodorsal es mucho más amplio que el margen anterodorsal y corre casi paralelo al margen ventral; el margen ventral es ligeramente curvo, siendo mayor la curva en las partes que se aproximan al margen posteroventral y anterodorsal. Placa charnelar con una carina interna delgada y paralela al margen dorsal. No presentan lúnula, ni escudo. La ornamentación es comarginal, con líneas muy delgadas y poco espaciadas entre ellas.
Material examinado. Once moldes externos con las valvas articuladas y cerradas (FCMP 886-896). De éstos, los ejemplares FCMP 895 y FCMP 896 presentan un desgaste que impide medir sus dimensiones.
Medidas. (Tabla 6).
Distribución. El registro del género Cardiomorpha abarca desde el Emsiano Tardío (Devónico) hasta el Wuchiapingiano (Pérmico Tardío). En el Reino Unido ha sido reportado para el Asbiano (Moore, 1958) y Brigantiano (Wolfenden, 1958); en China se refirió para el Eifeliano (Zhang y Pojeta, 1986) y el Wuchiapingiano (Zhao y Cheng, 1988); en India se reportó para el Sakmariano (Reed, 1932); en la Antártida se ha referido para el Serpukhoviano-Bashkiriano (Kelly et al., 2001); en Brasil se reportó para el Emsiano (Melo, 1988) y el Eifeliano-Frasniano (Morsch, 1986); en Estados Unidos se refirió para el Serpukhoviano de Virginia del Oeste (Hoare, 1993), el Desmoinesiano de Missouri (Hoare, 1961), Pensilvánico de Utah (Chamberlain, 1981), Misisípico de Alaska (Buddington y Chapin, 1929), Roadiano de Idaho (Girty, 1910a) y el Merameciano de Nevada (Brew y Gordon, 1971) y de Arkansas (Branson, 1958).
Discusión. El material examinado presenta similitud con los géneros Edmondia y Cardiomorpha debido a que ambos géneros tienen valvas ovaladas o subcuadradas y umbos prosogiros. Sin embargo, se diferencian en que los umbos de Edmondia están más desplazados hacia el centro ubicándose a un tercio desde el margen anterodorsal hacia el margen posterodorsal, mientras que los umbos de Cardiomorpha son casi terminales. Se determinó que los ejemplares revisados pertenecen al género Cardiomorpha por presentar éste último carácter.
El material examinado presenta afinidad con el material estudiado por Girty (1910a), quien describió un ejemplar de Cardiomorpha sp. como una concha con forma transversalmente subelíptica, con un umbo no muy prominente y casi terminal, con extremos simétricamente redondeados, y una superficie marcada por muchas estrías tenues y finas.
Pojeta (1969), al revisar los bivalvos de Girty (1910b), los asignó con duda a la especie Cardiomorpha? inflata, cuyos caracteres son distintivos del género, pero se distingue del material examinado principalmente en que su margen posterior es subrectilíneo y fuertemente oblicuo, haciendo que las valvas se observen alargadas y oblicuas, carácter del que carecen los ejemplares examinados, pues éstos tienen el margen posterior redondeado y, aunque sus valvas están alargadas, no son oblicuas.
La especie Cardiomorpha missouriensis presentada en Hoare et al. (1979) tiene el margen posterior subtruncado, mientras que el margen posterior del material revisado es redondeado, por lo tanto, no son equiparables. En ese trabajo se muestra la charnela del ejemplar que allí se determina; ésta tiene unas ninfas bien desarrolladas y una placa charnelar que presenta una delgada cresta interna que corre paralela a ella. En el material que se revisó hay unos ejemplares en los que las valvas están desplazadas; allí se expone una parte de la charnela cuya forma es similar a la que aparece en Hoare et al. (1979), aunque no se observa la cresta interna, lo cual se puede explicar por el hecho de que se rellenó de sedimento durante su fosilización y se perdió el carácter.
Al comparar el material revisado con ejemplares de Cardiomorpha obovata presentados en Hoare (1993), se observan grandes diferencias, pues los ejemplares de Hoare tienen los umbos muy desplazados hacia el centro, y el mismo autor los describe como umbos prosogiros, localizados aproximadamente a un cuarto de longitud desde el margen anterior; al observar las figuras, los ejemplares tienen más afinidad con formas parecidas a Edmondia. En el mismo trabajo aparecen dos ejemplares determinados como Cardiomorpha sp., los cuales sí presentan características afines al género, pero son distintos al material revisado porque sus valvas son más alargadas y con una forma más subcuadrada, además tienen un margen anterior liso y convexo hacia su margen anteroventral, caracteres que no presenta el material examinado.
Localidad. Camino Olinalá-El Progreso, localidad 2.
Figura 10. 1-11: ?Cardiomorpha sp.; 1, valva izquierda, 2, valva derecha, 3, vista dorsal, 4, vista anterior, ejemplar FCMP 888; 5, valva izquierda, 6, valva derecha, 7, vista dorsal, 8, vista anterior, ejemplar FCMP 894; 9, valva derecha, ejemplar FCMP 896; 10, valva izquierda, ejemplar FCMP 892; 11, valva derecha, ejemplar FCMP 893.
8. Conclusiones
Los bivalvos estudiados provienen de la lutita media de la Formación Olinalá, que corresponde a la unidad 4 de Corona-Esquivel (1981 [1983]) y González-Arreola et al. (1994), y a la parte lutítica de la secuencia SD-2 de Juárez-Arriaga (2006).
Se identificaron cuatro especies: Nuculavus levatiformis, Palaeoneilo mcchesneyana, Septimyalina burmai y Edmondia phosphatica. Tres especies sólo pudieron ser determinadas a nivel de género: Posidoniella sp., Bakevellia sp. y Leptodesma (Leptodesma) sp. Una especie se relacionó con duda al género Cardiomorpha.
Las especies estudiadas confirman la edad Pérmico medio (Guadalupiano) de la Formación Olinalá y sugieren una posible edad Roadiano para el nivel estudiado.
Los géneros encontrados representan tres modos de vida principales: Nuculavus, Palaeoneilo, Edmondia y Cardiomorpha incluyen especies que fueron organismos móviles facultativos y formaron parte de la infauna; Posidoniella, Septimyalina y Leptodesma incluyen especies que vivían adheridas a diferentes sustratos formando parte de la epifauna y Bakevellia es un género con especies semi-infaunales. Los cuatro últimos géneros han sido asociados comúnmente a aguas costeras y salobres. Estas diferencias ecológicas y las características particulares de las concentraciones fósiles y de la roca portadora, indican que la lutita donde se hallaron los fósiles corresponde a un ambiente de plataforma de aguas poco profundas de baja energía, influenciado por la desembocadura de ríos.
La afinidad que tienen las especies de bivalvos estudiadas con las faunas de la parte central y centro-oeste de Norteamérica, indica una relación con las provincias Cordillerana y Laurencia Sur como parte del Reino Americano de Bambach (1990). De acuerdo a las provincias pérmicas de Yancey (1975) corresponden a las provincias Grandiana y Cordillerana, consideradas por Shen et al. (2009) dentro del Reino Paleoecuatorial. Esta afinidad representa la continuidad del mar somero que cubrió la región de Olinalá con el mar que se extendió en la parte centro-oeste del norte de América durante el Pérmico medio.
Agradecimientos
Los autores agradecen a la Universidad Nacional Autónoma de México su apoyo a esta investigación a través de los proyectos PAPIIT IN 213710 e IN215013; a Leonora Martín-Medrano por la elaboración de los moldes en silicón; a Daniel Navarro-Santillán, Edgar Juárez-Arriaga y Francisco Sour-Tovar por su gran ayuda en el trabajo de campo y a Héctor Hernández-Campos por la elaboración de las fotografías del material estudiado. También, se agradece a María del Carmen Perrilliat (Instituto de Geología, UNAM) y a Luiz Eduardo Anelli (Universidade de Sao Paulo, Brasil) su cuidadosa revisión y críticas constructivas para mejorar el manuscrito.
Referencias
Adams, H., Adams, A., 1858, The genera of recent Mollusca: arranged according to their organization: London, John Van Voorst, 3, 484 p.
Anelli, L.E., Rocha-Campos, A.C., Simões, M.G., 2002, Protobranch bivalves from the Piauí Formation (Middle Pennsylvanian), Parnaíba Basin, Brazil: Revista Española de Paleontología, 17, 165-176.
Anelli, L.E., Rocha-Campos, A.C., Simões, M.G., 2006, Pennsylvanian pteriomorphian bivalves from the Piauí Formation, Parnaíba basin, Brazil: Journal of Paleontology, 80(6), 1125-1141.
Bambach, R.K., 1990, Late Palaeozoic provinciality in the Marine Realm, en McKerrow, W.S., Scotese, C.R., (eds.), Palaeozoic Palaeogeography and Biogeography: Geological Society Memoir, 12, 307-323.
Bambach, R.K., Scotese, C.R., Ziegler, A.M., 1980, Before Pangea: The Geographies of the Paleozoic World: American Scientist, 68, 26-38.
Bieler, R., Carter, J.G., Coan, E.V., 2010, part. 2, Classification of Bivalve Families, en Bouchet, P., Rocroi, .J.-P., Bieler, R., Carter, J.G., Coan, E.V. (eds.), Nomenclator of bivalve families with a Classification of Bivalve Families: Malacologia, 52(2), 1-184.
Blodgett, R.B., Moore, T.E., Gray, F., 2002, Stratigraphy and Paleontology of Permian Lower Rocks of north Cananea, northern Sonora, Mexico: Journal of South American Earth Sciences, 15, 481-495.
Böse, E., 1919, The Permo-Carboniferous ammonoids of the Glass Mountains, west Texas, and their stratigraphic significance: University of Texas Bulletin, 1762, 1-241.
Boyd, D.W., Newell, N.D., 1979, Permian pelecypods of Tunisia: American Museum Novitates, 2686, 1-22.
Branson, C.C., 1948, Bibliographic index of Permian invertebrates: Geological Society of America Memoir, 26, 1-1049.
Branson, C.C., 1958, Two Mississippian species of Conocardium: Oklahoma Geology Notes, 18, 137-142.
Brew, D.A., Gordon, M., 1971, Mississippian stratigraphy of the Diamond Peak Area, Eureka County, Nevada: United States Geological Survey Professional Paper, 661, 1-84.
Buddington, A.F., Chapin, T., 1929, Geology and mineral deposits of southeastern Alaska: United States Geological Survey Bulletin, 800, 1-398.
Carter, J.G., Campbell, D.C., Campbell, M.R., 2000, Cladistic perspectives on early bivalve evolution, en Harper, E.M., Taylor, J.D., Crame, J.A. (eds.), The evolutionary biology of the Bivalvia: Geological Society Special Publication, 177, 1-494.
Chamberlain, A.K., 1981, Biostratigraphy of the Great Blue Formation: Bringham Young University Geology Studies, 28(3), 9-17.
Chernyshev, B.I., 1947, Dekayi Nuculidae Donets´kogo basenyu: Akademie Nauk Ukrains´koyi RSR (Kiev), Instytut Geologichnykh Nauk, Zbirnyk prats´z paleontologiyi ta Stratigrafiyi, 1, 3-19.
Chronic, H., 1952, Molluscan fauna from the Permian Kaibab Formation, Walnut Canyon, Arizona: Geological Society of America Bulletin, 63(2), 95-166.
Ciriacks, K.W., 1963, Permian and Eotriassic bivalves of the Middle Rockies: Bulletin of the American Museum of Natural History, 125(1), 1-100.
Clifton, R.L., 1942, Invertebrate faunas from the Blaine and the Dog Creek Formations of the Permian Leonard Series: Journal of Paleontology, 16(6), 685-699.
Cloud, P.E., 1944, Geology and paleontology of the Permian area northwest of Las Delicias, southwestern Coahuila, Mexico, Permian brachiopods: Geological Society of America Special Paper, 52, 49-69.
Conrad, T.A., 1842, Observations on the Silurian and Devonian systems of the United States, with descriptions of new organic remains: Journal of the Academy of Natural Sciences of Philadelphia, 8, 228-280.
Cooper, G.A., 1953, Sponges, Brachiopoda, Pelecypoda, and Scaphopoda. Permian fauna at El Antimonio, western Sonora, Mexico: Smithsonian Miscellaneous Collections, 119(2), 21-80.
Cooper, G.A., Grant, R.E., 1972, Permian brachiopods of west Texas, I: Smithsonian Contributions to Paleobiology, 14, 231 p.
Corona-Esquivel, R., 1981 (1983), Estratigrafía de la región de Olinalá-Tecocoyunca, noreste del estado de Guerrero: Revista del Instituto de Geología, 5(1), 17-24.
Corona-Esquivel, R., 1985, Geología de la región comprendida entre Olinalá y Huamuxtitlán, Estado de Guerrero: Ciudad de México, Universidad Nacional Autónoma de México, tesis de licenciatura, 99 p.
Cox, L.R., Newell, N.D., Boyd, D.W., Branson, C.C, Casey, R., Chavan, A., Coogan, A.H., Dechaseaux, C., Fleming, C.A., Haas, F., Hertlein, L.G., Kauffman, E.G., Keen, A.M., LaRocque, A., McAlester, A.L., Moore, R.C., Nuttall, C.P., Perkins, B.F., Puri, H.S., Smith, L.A., Sooth-Ryen, T., Stenzel, H.B., Trueman, E.R., Turner, R.D., Weir, J., 1969, Treatise on Invertebrate Paleontology, part N Mollusca, 6. Bivalvia: Lawrence, The Geological Society of America and The University of Kansas Press, 952 p.
Dall, W.H., 1889, On the hinge of pelecypodes and its development, with an attempt toward a better subdivision of the group: American Journal of Science, 38, 445-462.
Dickins, J.M., 1999, Mid-Permian (Kubergandian-Murgabian) bivalves from the Khuff Formation, Oman: implications for world events and correlation: Rivista Italiana di Paleontologia e Stratigrafia, 105(1), 23-36.
Erben, H.K., 1956, El Jurásico Inferior de México y sus amonitas: XX Congreso Geológico Internacional, México, Universidad Nacional Autónoma de México, Instituto de Geología, 393 p.
Esquivel-Macías, C., 1996, Invertebrados del Paleozoico Tardío de las regiones de Olinalá, Gro. y Patlanoaya, Pue.: Ciudad de México, Universidad Nacional Autónoma de México, tesis de maestría, 82 p.
Esquivel-Macías, C., 2000, Braquiópodos y crinoides del Paleozoico Tardío de las Formaciones Olinalá, Gro.; Patlanoaya y Cuxtepeque, Pue.: Ciudad de México, Universidad Nacional Autónoma de México, tesis de doctorado, 124 p.
Esquivel-Macías, C., Solís-Marín, F., Buitrón-Sánchez, B.E., 2004, Nuevos registros de placas columnares de crinoideos (Echinodermata, Crinoidea) del Paleozoico Superior de México, algunas implicaciones paleobiogeográficas y paleoambientales: Coloquios de Paleontología, 54, 15-23.
Fang, Z.J., 1987, Bivalves from the Upper part of the Permian in southern Hunan, China: Collection of Postgraduate Theses of the Nanjing Institute of Geology and Palaeontology, Academia Sinica, 1, 349-411.
Flores de Dios, A., 1986, Estratigrafía de la Montaña de Guerrero entre los límites con Puebla y Oaxaca: Ciudad de México, Instituto Politécnico Nacional, tesis de licenciatura, 85 p.
Flores de Dios, A., Buitrón, B.E., 1982, Revisión y aportes a la estratigrafía de la Montaña de Guerrero: Universidad Autónoma de Guerrero, Serie Técnico-Científica, 12, 28 p.
Flores de Dios, A., Vachard, D., Buitrón, B.E., 2000, La cubierta sedimentaria pérmica superior (formaciones Olinalá, Ihualtepec y Yododeñe) de los terrenos Zapoteco y Mixteco, evolución sedimentológica e interpretaciones paleogeográficas (resumen), en II Reunión Nacional de Ciencias de la Tierra, Puerto Vallarta, Jalisco, México: Unión Geofísica Mexicana, GEOS, Resúmenes y Programa, 20(3), 21-22.
Frech, F., 1891, Die devonischen Aviculiden Deutschlands, ein Beitrag zur Systematik und Stammesgeschichte der Zweischaler: Abhandlungen zur geologischen Specialkarte von Preussen und Thüringischen Staaten, 9, 253 p.
García-Díaz, J.L., Marc, T., Campa-Uranga, M.F., Henrriette, L., 2000, Las Ignimbritas de Olinalá, Guerrero (Sierra Madre del Sur): Testigos de un margen activo continental Pacífico durante el Triásico?-Jurásico Medio, en II Reunión Nacional de Ciencias de la Tierra, Puerto Vallarta, Jal., Unión Geofísica Mexicana, GEOS, Resúmenes y Programa, 20(3), 329-330.
Garibay-Romero, L.M., 2000, Nuevos amonitas pérmicos de la Formación Olinalá-Los Arcos, Olinalá, Gro., México (resumen), en II Reunión Nacional de Ciencias de la Tierra, Puerto Vallarta, Jal., Unión Geofísica Mexicana, GEOS, Resúmenes y programa, 20(3), 299.
Geinitz, H.B., 1866, Carbonformation und Dyas in Nebraska: Kaiserliche deutsche Akademie Naturforscher zu Halle: Nova Acta Leopoldina, Dresden, Verhandlung, 33, 91 p.
Girty, G.H., 1908, The Guadalupian fauna: United States Geological Survey Professional Paper, 58, 1-651.
Girty, G.H., 1910a, The fauna of the phosphate beds of the Park City Formation in Idaho, Wyoming and Utah: United States Geological Survey Bulletin, 436, 82 p.
Girty, G.H, 1910b, New genera and species of Carboniferous fossils from the Fayetteville shale of Arkansas: Annals of the New York Academy of Sciences, 20(3) part 2, 189-238.
Goldfuss, G.A., 1833-1840, Petrefacta Germaniae: Arus and Company, 312 p.
González-Arreola, C., Villaseñor-Martínez, A.B., Corona-Esquivel, R., 1994, Permian fauna of the Los Arcos Formation, Municipality of Olinalá, State of Guerrero, Mexico: Revista Mexicana de Ciencias Geológicas, 11(2), 214-221.
Gray, J.E., 1824, Conchological observations, being an attempt to fix the study of conchology on a firm basis: The Zoological Journal, 1(2), 204-223.
Gray, J.E., 1847, A list of the genera of Recent Mollusca, their synonyms and types: Proceedings of the Zoological Society of London, 15, 129-219.
Hall, J., 1883, Lamellibranchiata, plates and explanations: New York Geological Survey Paleontology, 5(1), 20 p.
Hall, J., 1885, Lamellibranchiata II, descriptions and figures of the Dimyaria of the Upper Heidelberg, Hamilton, Portage and Chemung groups: New York Geological Survey Paleontology, 5, pt. 1, 269-562.
Hall, J., Whitfield, R.P., 1869, Preliminary notice of the lamellibranchiate shells of the upper Heidelberg, Hamilton and Chemung groups, with others from the Waverly sandstones, pt. 2, 80 p.
Harker, P., Thorsteinsson, R., 1960, Permian rocks and faunas of Grinnell Peninsula, Arctic Archipelago: Geological Survey of Canada Memoir, 309, 89 p.
Hind, W., 1896-1900, A monograph of the British Carboniferous Lamellibranchiata, 1. (1898): 1 (3): Nuculidae, Trigonidae, Unionidae, Edmondidae: Monograph of the Palaeontographical Society, 52, 209-276.
Hoare, R.D., 1961, Desmoinesian Brachiopoda and Mollusca from southwest Missouri: Missouri University Studies, 36, 262 p.
Hoare, R.D., 1993, Mississippian (Chesterian) bivalves from the Pennsylvanian Stratotype area in West Virginia and Virginia: Journal of Paleontology, 67(3), 374-396.
Hoare, R.D., Sturgeon, M.T., Kindt, E.A., 1978, New Pennsylvanian bivalves and occurrences of Pseudoconocardium from Ohio and West Virginia: Journal of Paleontology, 52(5), 1023-1036.
Hoare, R.D., Sturgeon, M.T., Kindt, E.A., 1979, Pennsylvanian marine Bivalvia and Rostroconchia of Ohio: Ohio Geological Survey Bulletin, 67, 77 p.
Hoare, R.D., Heaney III, M.J., Mapes, R.H., 1989, Bivalves (Mollusca) from the Imo Formation (Mississippian, Chesterian) of North-Central Arkansas: Journal of Paleontology, 63(5), 582-603.
Isaacson, P., Dutro, T., 1999, Lower Carboniferous brachiopods from Sierra de Almeida, northern Chile: Journal of Paleontology, 73(4), 625-633.
Juárez-Arriaga, E., 2006, Marco de Estratigrafía de Secuencias para la Sucesión Sedimentaria Terrígeno-Carbonatada del Paleozoico Superior del área de Olinalá, Guerrero, México: México, D.F., Universidad Nacional Autónoma de México, tesis de maestría, 93 p.
Kelly, S.R., Doubleday, P.A., Brunton, C.H., Dickins, J.M., Sevastopulo, G.D., Taylor, P.D., 2001, First Carboniferous and Permian marine macrofaunas from Antarctica and their tectonic implications: Journal of the Geological Society, 158(2), 219-232.
Kidwell, S.M., Fürsich, F.T., Aigner, T., 1986, Conceptual Framework for the Analysis and Classification of Fossil Concentrations: Palaios, 1, 228-238.
King, W., 1844, On a new genus of Palaeozoic shells: Annals and Magazine of Natural History, 14(92), 313-317.
King, W., 1848, A catalogue of the organic remains of the Permian rocks of Northumberland and Durham, Newcastle upon Tyne.
King, W., 1850, A monograph of the Permian fossils of England: Palaeontographical Society, XXXVIII, 258 p.
Koninck, L.G., De, 1841-1844, Description des animaux fossiles qui se trouvent dans le terrain carbonifère de Belgique. H. Dessain, Liège, 650 p.
Koninck, L.G., De, 1885, Marine Fauna of the Early Permian (Wolfcampian) Robledo Mountains Member, Hueco Formation, Southern Robledo Mountains, New Mexico, en Lucas, S.G., Heckert, A.B. (eds.), Early Permian footprints and facies: New Mexico Museum of Natural History and Science Bulletin, 6, 63-90.
Kues, B.S., 1995, Marine Fauna of the Early Permian (Wolfcampian) Robledo Mountains Member, Hueco Formation, Southern Robledo Mountains, New Mexico, en Lucas, S.G., Heckert, A.B. (eds.), Early Permian footprints and facies: New Mexico Museum of Natural History and Science Bulletin, 6, 63-90.
Lee, T., Girty, G.H., 1909, The Manzano Group of the Rio Grande Valley: United States Geological Survey Bulletin, 389, 120 p.
Linnaeus, C., 1758, Systema Naturae per regna tria naturae. Secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis, Tomus I.: Stockholm, Laurentii Salvii, Editio decima, reformata, 824 p.
Logan, A., 1967, The Permian bivalvia of northern England: Palaeontographical Society Monographs, 121, 72 p.
Marcou, J., 1858, Geology of North America with two reports on the Prairies of Arkansas and Texas, the Rocky Mountains of New Mexico, and the Sierra Nevada of California: Zürcher and Furrer, 144 p.
Mather, K.F., 1915, The Fauna of the Morrow Group of Arkansas and Oklahoma: Denison University Scientific Laboratories Bulletin, 18, 59-284.
Mayou, T.V., 1967, Paleontology of the Permian Loray Formation in White Pine County, Nevada: Brigham Young University Research Studies, Geology Series, 14, 101-122.
McAlester, A.L., 1969, Family Praenuculidae, en Cox, L.R., Newell, N.D., Branson, C.C., Casey, R., Chavan, A., Coogan, A.H., Dechaseaux, C., Fleming, C.A., Haas, F., Hertlein, L.G., Keen, A.M., LaRocque, A., McAlester, A.L., Perkins, B.F., Puri, H.S., Smith, L.A., Soot-Ryen, T., Stenzel, H.B., Turner, R.D., Weir, J., Treatise on Invertebrate Paleontology, Part N, Mollusca 6 (Bivalvia), N229, University of Kansas Press and Geological Society of America, United States of America.
Meek, F.B., 1864, Remarks on the family Pteriidae (=Aviculidae) with descriptions of some new fossil genera: The American Journal of Science and Arts, ser. 2, 37, 212-220.
Meek, F.B., Hayden, F.V., 1858, Descriptions of new organic remains collected in Nebraska territory together with some remarks on the geology of the Black Hills and portions of the surrounding country: Proceedings of the Academy of Natural Sciences of Philadelphia, 8, 70-72.
Melo, J.H., 1988, The Malvinokaffric realm in the Devonian of Brazil, en McMillan, N.J., Embry, A.F., Glass, D.J., (eds.), Devonian of the World: Canadian Society of Petroleum Geologists, Calgary, Alberta, Canada, I, 669-703.
Miller, S.A., 1877, The American Palaeozoic fossils: (publicado por el autor), Cincinnati, Ohio, xv + 253 p.
Miller, S.A., 1889, North American geology and palaeontology, 664 p.
Miller, A.K., Furnish., W.M., 1940, Permian ammonoids of the Guadalupe Mountain region and adjacent area: Geological Society of America Special Paper, 26, 1-242.
Moore, D., 1958, The Yoredale Series of Upper Wensleydale and adjacent parts of North-West Yorkshire: Proceedings of the Yorkshire Geological Society, 31(2), 91-148.
Morningstar, H., 1922, Pottsville fauna of Ohio: Ohio Geological Survey Bulletin, 25, 312 p.
Morsch, S.M., 1986, Bivalves (Mollusca) na Formação Ponta Grossa (Bacia do Paraná-Devoniano): Revisão Sistemática: Anais da Academia Brasileira de Ciências, 58(3), 403-431.
Mudge, M.R., Yochelson, E.L., Douglas, R.C., Duncan, H., Strimple, H.L., Gordon, M., Dunkler, D.H., 1962, Stratigraphy and Paleontology of Uppermost Pennsylvanian and Lowermost Permian rocks in Kansas: United States Geological Survey Professional Paper, 323, 209 p.
Newell, N.D., 1942, Late Paleozoic Pelecypods: Mytilacea: University of Kansas Publications, 10(2), 80 p.
Newell, N.D., 1965, Classification of the Bivalvia: American Museum Novitates, 2206, 25 p.
Newell, N.D., Chronic, J., Roberts, T.G., 1953, Upper Paleozoic of Peru: The Geological Society of America Memoir, 58, 276 p.
Ortega-Gutiérrez, F., Elías-Herrera, M., Reyes-Salas, M., Macías-Romo, C., López, R., 1999, Late Ordovician-Early Silurian continental collisional orogeny in southern Mexico and its bearing on Gondwana-Laurentia connections: Geology, 27, 719-722.
Phillips, J., 1836, Illustrations of the geology of Yorkshire, part. 2, the Mountain Limestone District, London, John Murray, 253 p.
Pojeta Jr., J., 1969, Revision of some of Girty’s invertebrate fossils from the Fayetteville shale (Mississippian) of Arkansas and Oklahoma-Pelecypods: United States Geological Survey Professional Paper, 606-C, 15-24.
Posenato, R., Pelikán, P., Hips, K., 2005, Bivalves and Brachiopods near the Permian-Triassic boundary from the Bükk Mountains (Bálvány-North Section, northern Hungary): Rivista Italiana di Paleontología e Stratigrafia, 111(2), 215-232.
Quiroz-Barroso, S.A., Sour-Tovar, F., Centeno-García, E., 2012, Gasterópodos y bivalvos cisularianos (Pérmico inferior) de Otlamalacatla, Hidalgo, México: Revista Mexicana de Ciencias Geológicas, 29(1), 158-178.
Reed, F.R., 1932, New fossils from the agglomerate Slate of Kashmir: Palaeontological Indica, Memoirs of the Geological Survey of India, 20(1), 79 p.
Runnegar, B., Newell, N.D., 1974, Edmondia and the Edmondiacea Shallow-Burrowing Paleozoic Pelecypods: American Museum Novitates, 2533, 19 p.
Sampelayo, P., Sampelayo, A., 1947, Fauna Carbonífera de Villablino (León): Notas y Comunicaciones del Instituto Geológico y Minero de España, 17, 3-24.
Servicio Geológico Mexicano (SGM), 2006, Léxico Estratigráfico de México http://www.sgm.gob.mx, consultado el 10 de octubre de 2012.
Shen, S.Z., Xie, J.F., Zhang, H., Shi, G.R., 2009, Roadian–Wordian (Guadalupian, Middle Permian) global palaeobiogeography of brachiopods: Global and Planetary Change, 65(3-4), 166-181.
Silva-Pineda, A., Buitrón, B.E., Flores de Dios, A., 1998, Biota (continental y marina) del Pérmico de la región de Olinalá, estado de Guerrero (resumen), en 1ª Reunión Nacional de Ciencias de la Tierra, Puerto Vallarta, Jalisco, México: GEOS, Unión Geofísica Mexicana, Resúmenes y Programa, 55 p.
Silva-Pineda, A., Buitrón, B.E., Flores de Dios, A., 2000, Presencia de coníferas en el Pérmico (Leonardiano) de Calnalli, Hidalgo; Patlanoaya, Puebla; y Olinalá, Guerrero (resumen), en 2ª Reunión Nacional de Ciencias de la Tierra, Puerto Vallarta, Jalisco, México: GEOS, Unión Geofísica Mexicana, Resúmenes y Programa, 20(3), 298-299.
Sour-Tovar, F., Quiroz-Barroso, S.A., Applegate, S.P., 2000, Presence of Helicoprion (Chondrichthyes, Elasmobranchii) in the Permian Patlanoaya Formation, Puebla, Mexico: Journal of Paleontology, 74(2), 363-366.
Sour-Tovar, F., Juárez-Arriaga, E., Murillo-Muñetón, G., 2009, Nuevos braquiópodos para el Pérmico del área de Olinalá, Guerrero, México: XI Congreso Nacional de Paleontología, 73.
Sowerby, J., 1823-1825, The mineral conchology of Great Britain; or coloured figures and descriptions of those remains of testaceous animals or shells, which have been preserved at various times and depths in the earth: London, Vol. V, 168 p.
Sowerby, J., 1829, The mineral conchology of Great Britain; or coloured figures and descriptions of those remains of testaceous animals or shells, which have been preserved at various times and depths in the earth: London, Vol. VI, 250 p.
Sterren, A.F., 2000, Moluscos bivalvos en la Formación Río del Peñón, Carbonífero tardío-Pérmico temprano, provincia de La Rioja: Ameghiniana, 37(4), 421-438.
Sterren, A.F., 2004, Bivalvos pérmicos de la Formación Tupe en la quebrada de La Herradura, provincia de San Juan: Ameghiniana, 41(1), 57-74.
Vachard, D., Oviedo, A., Flores de Dios, A., Malpica, R., Brunner, P., Guerrero, M., Buitrón, B.E., 1993, Barranca d’Olinalá (Guerrero): Une coupe de réference pour le Permien du Mexique central; étude préliminaire: Annales de la Société Géologique du Nord, 2, 155-162.
Vachard, D., Flores de Dios, A., Buitrón, B.E., 2004, Guadalupian and Lopingian (Middle and Late Permian) deposits from Mexico and Guatemala, a review with new data: Geobios, 37, 99-115.
Walcott, Ch.D., 1884, Paleontology of the Eureka District: United States Geological Survey Monograph, 8, 298 p.
Weihong, H., Feng, Q., Weldon, E.A., Gu, S., Meng, Y., Zhang, F., Wu, S., 2007, A Late Permian to Early Triassic Bivalve fauna from the Dongpan Section, Southern Guangxi, South China: Journal of Paleontology, 81(5), 1009-1019.
Winters, S.S., 1963, Supaí Formation (Permian) of eastern Arizona: Geological Society of America Memoir, 89, 99 p.
Wolfenden, E.B., 1958, Paleoecology of the Carboniferous reef complex and shelf limestones in the northwest Derbyshire, England: Bulletin of the Geological Society of America, 69, 871-898.
Woodward, S.P., 1854, A manual of the Mollusca being a treatise on recent and fossil shells: London, Crosby Lockwood and Co., 518 p.
Yancey, T.E., 1969, A molluscan faunal assemblage from the Arcturus Formation, Nevada: PaleoBios, 8, 20 p.
Yancey, T.E., 1975, Permian marine biotic provinces in North America: Journal of Paleontology, 49(4), 758-766.
Yancey, T.E., 1978, Brachiopods and Molluscs of the Lower Permian Arcturus Group, Nevada and Utah, part 1: brachiopods, scaphopods, rostroconchs, and bivalves: Bulletins of American Paleontology, 74(303), 257-367.
Yancey, T.E., Stevens, C.H., 1981, Early Permian fossil communities in northeastern Nevada and northwestern Utah, en Gray, J., Boucot, A.J., Berry, W.B.N., (eds.), Communities of the Past: Hutchinson Ross Publishing Co., Stroudsburg, Pennsylvania, 243-268.
Yochelson, E.L., 1956, Permian Gastropoda of the southwestern United States. 1. Euomphalacea, Trochonematacea, Pesudophoracea, Anomphalacea, Craspedostomatacea, and Platyceratacea: Bulletin of the American Museum of Natural History, 110(3):173-276.
Zhang, R., Pojeta, J., 1986, Devonian sections in Guangxi from which pelecypods were collected. Devonian rocks and Lower and Middle pelecypods of Guangxi, China and the Traverse Group of Michigan: United States Geological Survey Professional Paper, 1394-E, 33-43.
Zhao, J.M., Cheng, Z.X., 1988, A tabulate coral from the Lower Permian Toeling Formation at Douling of Changning County, Hunan, China: Acta Palaeontologica Sinica, 27, 650-652.
Ziegler, A.M., Hulver, M.L., Rowley, D.B., 1997, Permian world topography and climate, en Martini, I.P., (ed.), Late glacial and postglacial environmental changes: Quaternary, Carboniferous-Permian and Proterozoic: Oxford University, 111-146.
Manuscrito recibido: Junio 4, 2013.
Manuscrito corregido recibido: Agosto 4, 2013.
Manuscrito aceptado: Agosto 7, 2013.