
|
BOLETÍN DE LA SOCIEDAD GEOLÓGICA MEXICANA http://dx.doi.org/10.18268/BSGM2009v61n2a15 |
 |
Contribución al conocimiento de los rinocerontes fósiles de la Cuenca de Tecolotlán, en el estado de Jalisco, México
Contribution to knowledge of fossil rhinoceros of the Tecolotlán Basin in the state of Jalisco, Mexico
Gerardo Carbot–Chanona1,*, Javier Juárez–Woo2, José Rubén Guzmán–Gutiérrez3
1 Museo de Paleontología Eliseo Palacios Aguilera, Instituto de Historia Natural. Calzada de Los Hombres Ilustres s/n, C. P. 29000. Tuxtla Gutiérrez, Chiapas, México.
2 Museo de Paleontología de Guadalajara Federico A. Solórzano Barreto. Dr. R. Michel 520, C. P. 44100, Guadalajara, Jalisco, México.
3 Departamento de Paleontología, Desarrollo Turístico El Caracol, SECTURE, Av. Perseo s/n. Aguascalientes, Aguascalientes, México.
* This email address is being protected from spambots. You need JavaScript enabled to view it..
Resumen
Se describe el material de Teleoceras hicksi (Perissodactyla, Rhinocerotidae), depositado en el Museo de Paleontología de Guadalajara y en el Museo Regional de Guadalajara, proveniente de los sedimentos fluviolacustres de la Cuenca de Tecolotlán, Jalisco, México. Los restos examinados consisten en un cráneo incompleto con las series P2–M3 y una mandíbula asociada con ambos I2 y las series P3–M3, parte anterior de una mandíbula aislada con el I2 izquierdo y ambas series P3–M3, una mandíbula aislada con la serie P3–M3 pero sin sínfisis, molares superiores aislados, dos magno derechos, un metacarpo III izquierdo y la primera falange del dígito III derecho. Se observaron variaciones morfológicas en el material examinado que refleja diferencias de edad y/o sexo entre los individuos. Teleoceras hicksi se encuentra en la Cuenca de Tecolotán asociado con Dinohippus mexicanus, Nannippus aztecus, Neohipparion eurystyle, Astrohippus stockii y Borophagus secundus, por lo que la edad de la fauna corresponde al Henfiliano tardío.
Palabras clave: Cuenca de Tecolotlán, Jalisco, Rhinocerotidae, Teleoceras hicksi, Henfiliano tardío.
Abstract
Remains of Teleoceras hicksi (Perissodactyla, Rhinocerotidae), housed at the "Museo de Paleontologia "in Guadalajara and the "Museo Regional de Guadalajara", recovered from the fluvial lacustrine sediments in the Tecolotlán Basin are described. The studied material includes a partial skull bearing P2–M3 and one lower jaw preserving I2 and P3–M3, the anterior part of an isolated lower jaw with left I2 and P3–M3, one isolated lower jaw with P3–M3 dental series in wich the symphyseal region is missing, also isolated molars, two magnus, one left metacarpus III, and one first phalange of the right digit III. Morphological differences were observed in the material examined attributed to differences in age and/or sex of the individuals. Teleoceras hicksi was found associated with Dinohippus mexicanus, Nannippus aztecus, Neohipparion eurystyle, Astrohippus stockii and Borophagus secundus, suggesting a late Hemphillian age for thefaunal association.
Keywords: Tecolotlán Basin, Jalisco, Rhinocerotidae, Teleoceras hicksi, late Hemphillian.
1. Introducción
Los rinocerontes fósiles son conocidos en México desde finales del siglo XIX, cuando E.D. Cope (1884) revisó los ejemplares fósiles de vertebrados del antiguo Museo Nacional de México, mencionando la mitad de la rama mandibular de Aphelops? fossiger (= Teleoceras fossiger) procedente del Valle de Toluca, Estado de México, la cual ahora está depositada en el Museo del Instituto de Geología de la Universidad Nacional Autónoma de México. A partir de ahí, se han registrado restos de Teleoceras en localidades miocénicas del norte, centro y sureste del país. Por ejemplo, T. fossiger ha sido reportado de localidades del Mioceno tardío de Michoacán (Brunet, 1968), Sinaloa (Maldonado–Köerdell, 1954) y Guanajuato (Carranza–Castañeda y Ferrusquía–Villafranca, 1978; Carranza–Castañeda, 1989), mientras que restos asignados a Teleoceras sp. provienen de localidades en Chihuahua, Jalisco, Guanajuato, Hidalgo, Morelos, Estado de México y Chiapas (Brunet, 1968; Lance, 1950; Lindsay, 1984;Ferrusquia–Villafranca, 1990; Castillo–Cerón et al., 1996; Miller y Carranza–Castañeda, 2002; Carbot–Chanona, 2008). Freudenberg (1921, 1922) incluyó a Teleoceras felicis como parte de la fauna de Tehuichila, Hidalgo, pero no mencionó el material en que basó su asignación y a la fecha se desconoce su paradero, haciendo imposible su revisión. Dalquest y Mooser (1980) describieron a T. ocotensis como una nueva especie para la Fauna Local Rancho El Ocote, con base en un M3, sin embargo Carranza–Castañeda (1989) consideró que se trata de un ejemplar joven y sinonimizó a esta especie con T. fossiger. Prothero (2005) a su vez sinonimizó a T. ocotensis con T. hicksi, y en consecuencia refirió todo el material de rinocerontes de la fauna Rancho El Ocote a esta especie. Es claro entonces, que a pesar de la extensa distribución geográfica de Teleoceras en México, muchos de los registros están basados en material fragmentario o pobremente conservado, lo que ha dificultado su asignación a nivel de especie; a excepción del material de rinocerontes proveniente de la Fauna del Rancho El Ocote, Guanajuato, el cual, hasta el momento es el mejor estudiado.
Los primeros trabajos sobre rinocerontes fósiles en el estado de Jalisco fueron hechos por Brunet (1968) quien reportó el género Teleoceras con base en un cráneo incompleto procedente de Tecolotlán (el mismo que se describe en este trabajo). Trabajos posteriores han mencionado la presencia de Teleoceras para la misma área (e.g. Rodríguez–Huerta y Guzmán–Gutiérrez, 1993; Miller y Carranza–Castañeda, 1998; Carranza–Castañeda, 2006), sin embargo, únicamente se menciona el material sin entrar en mayor detalle.
Según Brunet (1968), los ejemplares descritos en este trabajo fueron recolectados por pobladores de la región en los alrededores del poblado de Santa María, cerca de Tecolotlán, Jalisco, aproximadamente a 112 km al suroeste de la ciudad de Guadalajara (Figura 1) y posteriormente fueron donados a Federico Solórzano Barreto en el año 1960, quien depositó el material en el Museo de Paleontología de Guadalajara "Federico A. Solórzano Barreto" y en el Museo Regional de Guadalajara. Estudios posteriores señalan que todo el material de rinocerontes proviene de los sedimentos fluviolacustres de la Cuenca de Tecolotlán.
2. Geología y paleontología de la Cuenca de Tecolotlán
La Cuenca de Tecolotlán se sitúa en la Sierra Madre Occidental dentro de la Faja Volcánica Transmexicana, a 100 km al suroeste de Guadalajara; tiene un área de 18 km por 10 km y se orienta de norte a sur (Miller y Carranza–Castañeda, 1998). Carranza–Castañeda (2006) describió la secuencia fluviolacustre de la Cuenca de Tecolotlán, diferenciando dos secciones que en total miden alrededor de 80 m (Figura 2). La sección inferior de la secuencia está compuesta por estratos de arcilla, arena arcillosa finogranuda y caliche; en la parte superior está presente una capa de ceniza volcánica que ha sido datada mediante 40Ar/39Ar en una edad de 4.89 ± 0.016 Ma (Kowallis et al., 1998 en Carranza–Castañeda, 2006). De esta sección se han recuperado restos de Crocodylus sp., Gopherus sp., Notolagus velox, Hypolagus sp., Spermophilus sp., Calomys sp., Agriotherium schneideri, Pseudaelurus cf. P. intrepidus, Machairodus cf. M. coloradensis, Canis ferox, Astrohippus stockii, Nannippus aztecus, Neohipparion eurystyle, Dinohippus mexicanus, Rhyncotherium cf. R. falconeri, Hemiauchenia vera, Megalonyx sp., Megatylopus cf. M. matthewi, Borophagus secundus (=Osteoborus cyonoides) y Teleoceras hicksi. La edad de la sección ha sido determinada como Henfiliano tardío con base en la asociación faunística (Miller y Carranza–Castañeda, 1998; Carranza–Castañeda, 2006) y puede correlacionarse con las localidades de Yepómera, Chihuahua (Lance, 1950; Lindsay, 1984), El Ocote y Rinconada, Guanajuato (Carranza–Castañeda y Ferrusquía–Villafranca, 1978; Dalquest y Mooser, 1980; Carranza–Castañeda, 1992) y Teocaltiche, Jalisco (Montellano–Ballesteros, 1997).
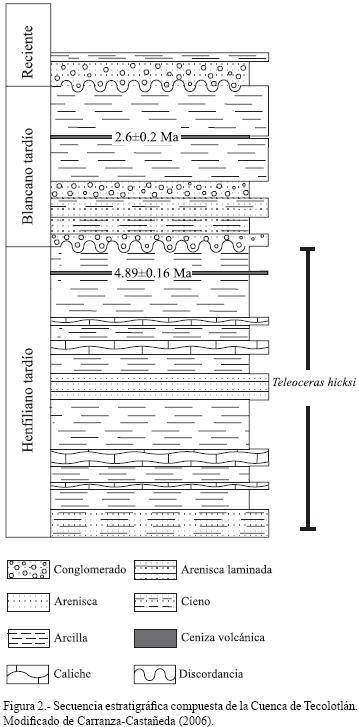
La sección superior de la secuencia está conformada por estratos de arena finogranuda y grava en la que también se observa una capa de ceniza volcánica datada en 2.6 ±0.2 Ma con el método 40Ar/39Ar (Kowallis et al., 1998 en Carranza–Castañeda, 2006). En esta sección se han recuperado restos de Glyptotherium sp., Neochoerus sp., Platygonus sp., cf. Plaina y Equus simplicidens que indican Blancano más tardío; sobre está parte se ubican discordantemente estratos conformados por arcilla y grava de edad reciente (Carranza–Castañeda, 2006).
3. Material y método
El material descrito en este trabajo fue comparado con el estudiado por Carranza–Castañeda (1989) proveniente de la fauna local Rancho El Ocote, Guanajuato, determinado como Teleoceras fossiger (= T. hicksi), así como con las descripciones e ilustraciones presentadas en los trabajos de Hatcher (1894), Osborn (1904) y Prothero (2005). La nomenclatura morfológica utilizada para la descripción está basada en Prothero (2005).
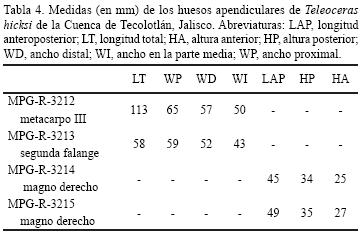
Las medidas se obtuvieron tomando la longitud y anchura de la superficie oclusal de molares y premolares superiores e inferiores que presentan distintos grados de desgaste. En el caso del cráneo y las mandíbulas se tomaron diferentes medidas, las cuales se detallan en las Tablas 1 y 2. Se describen cuatro elementos apendiculares y sus dimensiones se muestran en la Tabla 4. Todas las medidas se expresan en milímetros.
Las abreviaturas y siglas utilizadas en este trabajo son: I, incisivo; IGM, Instituto Geológico de México; PX, premolar superior; PX, premolar inferior; MX, molar superior; MX, molar inferior; mm, milímetros; MPG–R, Museo de Paleontología de Guadalajara–Referencia; MRG, Museo Regional de Guadalajara; UNAM, Universidad Nacional Autónoma de México.
4. Sistemática Paleontológica
Orden Perissodactyla Owen, 1848
Familia Rhinocerotidae Gray, 1821
Tribu Teleoceratini Hay, 1902
Género Teleoceras Hatcher, 1894
Teleoceras hicksi Cook, 1927
Especie tipo. Teleoceras major Hatcher, 1894. Clarendoniano temprano, Nebraska
Sinonimias. Paraphelops yumensis (Cook, 1930), Paraceras ponderensis (Cook, 1930), Teleoceras yumensis (Matthew, 1932), Teleoceras schultzi (Tanner, 1975), Teleoceras ocotensis (DalquestyMooser, 1980), Teleoceras fossiger (Carranza–Castañeda, 1989).
Diagnosis. Teleoceras de mediano tamaño (longitud del M1–M3= 150–165 mm, longitud del M3–M3= 155–180 mm) con cráneo más braquicefálico que T. fossiger. Nasales cortos y ligeramente levantados en muchos especímenes, pero no aplanados como en T. brachyrhinum. P2 ausente (Prothero, 2005).
4.1. Material referido
MRG–10–295030 cráneo (parte A) y mandíbula (parte B); MPG–R–0137, mandíbula con las series P3–M3 derecha e izquierda, pero sin la sínfisis; MPG–R–0248, parte anterior de la mandíbula con los I2, y las series P3–M3 derecha e izquierda; MPG–R–3200, M2 izquierdo; MPG–R–3201, M2 derecho; MPG–R–3202, M3 izquierdo; MPG–R–3203, M2 derecho; MPG–R–3204, M2 derecho; MPG–R–3205, M3 derecho; MPG–R–3207, M3 izquierdo; MPG–R–3208, fragmento del maxilar con el P2–M1 derechos; MPG–R–3209, fragmento del maxilar con el M1 y M2 izquierdos; MPG–R–3210, fragmento del maxilar con M2 y M3 izquierdos; MPG–R–3212, metacarpo III izquierdo; MPG–R–3213, primera falange del dígito III; MPG–R–3214, magno derecho; MPG–R–3215, magno derecho.
4.2. Descripción del material
MRG–10–295030–A consiste en la parte anterior de un cráneo bien conservado (Figura 3), en el cual toda la región occipital y los parietales no están presentes. Los arcos zigomáticos comienzan al nivel de M1, les falta la porción posterior, pero aun así puede apreciarse que son más estrechos anterior que posteriormente. Por encima de las órbitas se observan los procesos supraoccipitales bien desarrollados, lo que hace que los huesos frontales sean anchos y que se estrechen poco posteriormente. Premaxilares no fusionados, más largos que los nasales y conservan los alveolos de los I1. Los nasales son anchos pero se estrechan mucho hacia la parte anterior; los bordes laterales son redondeados, lo que hace que en sección transversal tengan forma de "U". Los molariformes están medianamente desgastados y se conservan las dos series dentarias completas, desde el P2 al M3, los cuales se describen individualmente a continuación.
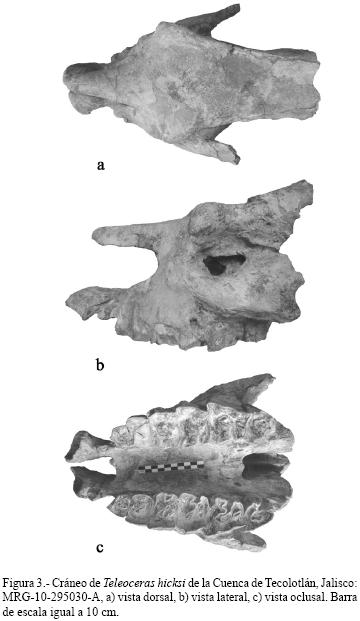
P2.– Premolar ligeramente desgastado. La prefoseta es alargada y está constreñida sobre su parte media;
postfoseta muy pequeña y de forma lenticular. El crochet y el anticrochet no se observan, posiblemente por el desgaste que presenta el diente. Valle medio cerrado por la fusión del metalofo y el protolofo. Cíngulo no presente. Parastilo reducido.
P3.– Premolar semicuadrado, con el parastilo muy pequeño; metalofo y protolofo fuertemente fusionados, cerrando el valle y formando la prefoseta, la cual es de mediano tamaño y alargada; postfoseta apenas visible. Crochet lobular y pequeño, anticrochet no visible. Cíngulo ligeramente visible en la pared del ectolofo.
P4.– Parastilo pequeño, ligeramente más proyectada labialmente que en el P3; protocono semicuadrado y ancho, ectolofo plano. Prefoseta alargada, postfoseta pequeña. Crochet muy pequeño, anticrochet no visible debido al desgaste del diente. Protolofo en contacto con el metalofo. Cíngulo lingual bien desarrollado.
M1.– Ectolofo ancho y convexo, parastilo medianamente desarrollado. Prefoseta alargada y abierta; postfoseta pequeña, constreñida en su parte media por el desarrollo del protolofo. Crochet no presente; anticrochet pequeño y casi en contacto con el protolofo. Parastilo medianamente desarrollado; cíngulo lingual apenas visible.
M2.– Prefoseta alargada y abierta sobre la cara lingual, postfoseta grande y de forma lenticular. Crochet redondeado y pequeño; anticrochet ancho y de forma lobular, de mayor tamaño que el crochet. Cíngulo visible por las caras anterior y posterior del diente. Metalofo y protolofo bien desarrollados.
M3.– Molar con el parastilo pequeño; protocono bien desarrollado. Prefoseta y mediofoseta constreñidas. Anticrochet muy pequeño, crochet más grande que el anticrochet, dirigido hacia él sin llegar a contactarlo. Cíngulo visible en el borde lingual del protolofo. Crista presente.
MRG–10–295030–B es una mandíbula en buen estado de preservación que conserva los dos I2 y las dos series dentarias P3–M3 (Figura 4a). Los cóndilos articulares están completos; los procesos coronoides son rectos y orientados ligeramente hacia delante, rasgo característico de los teleoceratinos. El diastema es corto; las series dentarias son paralelas pero las ramas mandibulares están en V cerrada. La sínfisis es ancha; los I2 son anchos, dirigidos lateralmente y en sección transversal son semiovalados. El M3 presenta un ligero cíngulo visible sólo por su parte labial. El foramen mentoniano está presente a la altura de la raíz del P4.
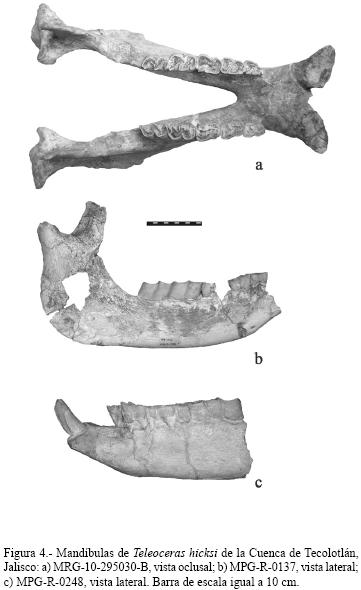
La morfología del cráneo y la mandíbula, el desgaste dental y la anchura del I2, indican que este ejemplar fue un macho adulto bien desarrollado.
MPG–R–0248. Mandíbula a la que le falta la mitad posterior de ambas ramas mandibulares, conservándose la serie P3–M3 derecha, y P4–M3 izquierda, así como el I2 izquierdo (Figura 4b). Las ramas mandibulares son más robustas que en MPG–R–0137 y el I2 es ligeramente más angosto transversalmente que el I2 de MRG–10–295030–B (Tabla 2); estas características indican que se trata de un macho adulto.
MPG–R–0137. Mandíbula casi completa a la cual le falta la sínfisis, del P3 en adelante, por lo que ambos I2 no están presentes (Figura 4c). A los procesos coronoides les falta la parte más proximal, sin embargo los cóndilos articulares están completos; el foramen mentoniano se encuentra ubicado ligeramente más abajo del comienzo de la raíz del P4. La dentición está conformada por P3 al M3 derechos e izquierdos, faltando únicamente la mitad anterior del M1 derecho. Las ramas mandibulares son ligeramente más gráciles que enMRG–10–295030–B y que en MPG–R–0248, lo que sugiere que se trata de un individuo juvenil.
4.2.1. Molariformes aislados
MPG–R–3209 es un fragmento del maxilar con el M1 y el M2 derechos (Figura 5a), de los cuales el segundo está muy fragmentado. M1 con un grado de desgaste mínimo, lo que sugiere que el individuo era juvenil. Parastilo bien desarrollado; la prefoseta es profunda, alargada y se encuentra abierta; postfoseta grande. Protolofo y protocono constreñidos. Metalofo angosto. El cíngulo es fuerte en toda la cara lingual. Ectolofo ancho y convexo, con una altura de 53.5 mm. Crochet pequeño; anticrochet ligeramente mayor pero más delgado. Crista reducida.
MPG–R–3200. M2 izquierdo con la prefoseta abierta; postfoseta pequeña. El crochet es de forma lobular, el anticrochet de forma similar pero ligeramente de mayor tamaño. El ectolofo es alto. Parastilo prominente. Metastilo, metacono y borde lingual del metalofo rotos. Hay cíngulos anterior y posterior (Figura 5b).
MPG–R–3201. M2 derecho con ectolofo reducido; protolofo prolongado. Protocono de forma semicuadrada por su parte lingual, con el esmalte formando una depresión. Parastilo y metastilo rotos. El cíngulo es visible en la parte anterior, posterior y lingual del diente. La postfoseta no se observa por el desgaste, pero la prefoseta es alargada. Crochet no visible por el desgaste del diente; anticrochet grande y de forma lobular (Figura 5c).
MPG–R–3203 es un M2 derecho, en el cual el borde lingual del protocono está fracturado. El parastilo es prominente y de forma lobular. Metastilo fracturado. El crochet es más pequeño que el anticrochet. Prefoseta y mediofoseta constreñidas. Postfoseta pequeña y cerrada. Borde del metalofo en forma de "U". Cíngulo no visible por la mala conservación del ejemplar (Figura 5d).
MPG–R–3204. M2 derecho con poco desgaste, que indica se trata de un animal joven. El ectolofo es ancho y convexo y tiene una atura de 75.2 mm. Prefoseta profunday abierta, ya que el valle no se ha fusionado. Postfoseta amplia. Crochet lobular y grande, anticrochet pequeño. Metalofo delgado y alargado El cíngulo no se observa (Figura 5e).
MPG–R–3210. Fragmento del maxilar que conserva el M2 y M3 izquierdos (Figura 5f); ambos molariformes presentan un desgaste marcado, lo que sugiere que pertenecieron a un animal adulto. En el M2 el parastilo es grande y de forma lobular. El crochet no es visible, contrariamente al anticrochet que es grande y ancho y casi contacta al metalofo, lo que hace que la prefoseta presente un estrechamiento en esa zona. La postfoseta es muy pequeña. El valle se encuentra abierto. El M3 tiene un protocono bien desarrollado. Parastilo y paracono pequeño. Metastilo prominente. Crochet muy reducido, debido tal vez al desgaste; anticrochet pequeño y de forma lobular. El valle medio está abierto desde la base. Ectolofo delgado. Cíngulo visible en todo el perímetro del molar.
MPG–R–3202. M3 izquierdo muy desgastado adherido a una porción del maxilar (Figura 5g). La prefoseta y postfoseta son profundas. El cíngulo es visible sólo en la parte lingual. Crochet y anticrochet de forma lobular y casi del mismo tamaño. Parastilo roto. Metastilo prominente. Protocono ancho.
MPG–R–3205 es un M3 derecho con el protocono bien desarrollado; parastilo y metastilo rotos. Prefoseta pequeña; postfoseta no visible debido a que esa parte del diente está perdida. Crochet pequeño, anticrochet de mayor tamaño. Cíngulo pequeño en el borde lingual del protolofo (Figura 5h).
MPG–R–3207. M3 izquierdo con ectolofo semiplano. El parastilo, al igual que el protocono, bien desarrollados. Metalofo y protolofo unidos. Cíngulo visible en toda la pared del ectolofo, metalofo y protolofo. Prefoseta y postfoseta unidas. Crochet lobular, anticrochet de igual forma pero ligeramente más ancho.
MPG–R–3208 es una porción maxilar que conserva del P2 al M1 en los que se observa un alto grado de desgaste (Figura 5i), que sugiere pertenecieron a un individuo viejo. El P2 sólo está representado por un fragmento del metalofo. En el P3 únicamente se observan remanentes del paracono, metacono, protocono y metalofo. P4 con parastilo prominente; metastilo reducido. Prefoseta apenas visible por el alto grado de desgaste. M1 muy gastado y sólo se diferencia el parastilo.
4.2.2. Restos apendiculares
MPG–R–3212 es un metacarpo III izquierdo bien conservado. Longitudinalmente es corto, pero robusto (Figura 6a), rasgo característico en Teleoceras. Los cóndilos lateral y medial son casi del mismo tamaño. La faceta articular para el unciforme es pequeña y forma una prominencia dirigida proximolateralmente. La faceta articular para el magno–trapezoide es plana y casi el doble de tamaño que la faceta articular para el unciforme. La faceta articular para el digito es amplia, tal como se observa en otros perisodáctilos.
MPG–R–3213 es una primera falange del digito III (Figura 6b y c). Es casi igual de larga que ancha, por lo que su forma es cuadrangular. El borde del extremo distal es ligeramente cóncavo, mientras que el proximal es casi plano. La faceta que articula con la segunda falange no es muy amplia.
MPG–R–3214 y MPG–R–3215 son magnos derechos (Figura 6d–g), ambos en relativamente buen estado de conservación y sin diferencias significativas en relación a los magnos de Teleoceras previamente descritos. En MPG–R–3215 la faceta articular para el metacarpo II es ligeramente más amplia que la de MPG–R–3 214 y el proceso posterior es menos prominente. En ambos especímenes el proceso posterior está bien desarrollado.
5. Discusión
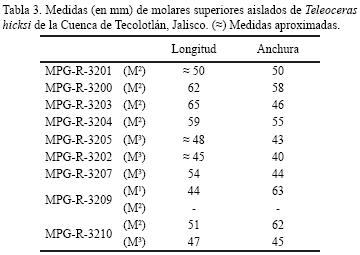
Prothero (2005) considera válidas tres especies para el Henfiliano de Norteamérica: T.fossiger (especie de mediano tamaño), presente en el Henfiliano temprano; T. guymonensis (especie de pequeño tamaño) y T. hicksi (especie de mediano tamaño), ambas presentes en el Henfiliano tardío. El material descrito en este trabajo es referido a Teleoceras hicksi con base en la siguiente combinación de caracteres, rinoceronte de mediano tamaño (longitud de las series M1–M3= 139 mm, M1–M3= 140 mm), nasales cortos que en sección transversal tienen forma de U y ligeramente orientados hacia arriba, y ausencia del P2. Aunque las dimensiones de las series dentales M1–M3 y M1–M3 de los especímenes provenientes de la Cuenca de Tecolotlán, son menores que los rangos de T. hicksi (M1–M3= 150–165 mm, M1–M3= 155–180 mm) y caen dentro de las medidas dadas para T. guymonensis (M1–M3= 159 mm; M1–M3= 140 mm), no se observó en ninguno de los M3 la costilla en la cara lingual del crochet (que forma un pequeño espolón lingual sobre la superficie de desgaste), característico en esta especie (Prothero, 2005). Claramente, el ejemplar MRG–10–295030 era un animal de talla menor a la media establecida para T. hicksi. Sin embargo, se descarta la posibilidad de que se trate de una nueva especie, pues la comparación de los ejemplares de la Cuenca de Tecolotlán y los procedentes de la fauna Rancho El Ocote, Guanajuato, determinados como Teleoceras fossiger (= T. hicksi), no mostró diferencias morfológicas ni de tamaño (Tabla 1) que indiquen se trate de especies distintas. No obstante, se observaron pequeñas diferencias en la morfología craneal y dental que son atribuidas a variación intraespecífica. Por ejemplo, el cráneo MRG–10–295030–A tiene similitudes con el cráneo IGM–8357 del Rancho El Ocote, en la forma y constitución de los arcos zigomáticos, apreciándose que en ambos especímenes son más estrechos en su porción anterior que la posterior, mientras que en el cráneo IGM–8360 los arcos zigomáticos son casi del mismo ancho en ambos extremos. Las tres mandíbulas descritas en este trabajo no presentan diferencias morfológicas notorias entre sí, ni con el material de Guanajuato, observándose sólo variaciones en el tamaño y constitución de las ramas mandibulares, así como en las dimensiones de los I2 y de las series P3–M3 (Tabla 2). En los molariformes la morfología y dimensiones de la superficie oclusal es aun más variada (Tabla 3). Esto se aprecia al observar el M2, MPG–R–3204, en donde el crochet es grande y el anticrochet pequeño, al contrario de los M2, MPG–R–3210 y MPG–R–3200, en los cuales el crochet no se observa o es muy pequeño y el anticrochet está muy desarrollado. Algo similar se observa con la forma de la postfoseta de los M1, la cual es constreñida en MRG–10–295030–A, de tal forma que casi se tocan sus paredes interiores por el desarrollo del protolofo y en el M1 MPG–R–3209 es ancha y de forma lenticular. Esta variabilidad sin duda alguna es consecuencia del diferente grado de desgaste presente en la superficie oclusal de cada molariforme, que a su vez es producto del uso, indicando que pertenecieron a individuos de diferente edad. Por el contrario, las diferencias en la constitución craneal y mandibular, pueden ser indicativas de dimorfismo sexual. Carranza–Castañeda (1989) también observó estas variaciones morfológicas tanto en el cráneo, como en los molariformes de los ejemplares que estudió, llegando a la conclusión de que se debían a las diferencias individuales que se presentan en una misma población, las cuales a su vez están influenciadas por la edad y el sexo de cada individuo. Lo anterior también ha sido documentado en Teleoceras fossiger, T. major, T. proterum y Aphelops malacorhinus (Osborn 1898; Mead, 2000; Mihlbachler, 2005). Aunado a las características morfológicas de los especímenes descritos, la edad establecida para la fauna asociada a los restos de rinocerontes de la Cuenca de Tecolotlán es un elemento más que sustenta la asignación del material como T. hicksi.
6. Conclusiones
La revisión del material de rinocerontes depositado en el Museo de Paleontología de Guadalajara y en el Museo Regional de Guadalajara, confirma la presencia del rinoceronte Teleoceras hicksi en los sedimentos fluviolacustres de la Cuenca de Tecolotlán, Jalisco. El estudio de los caracteres morfológicos, así como las medidas del material descrito en este trabajo mostraron diferencias mínimas en el cráneo, mandíbulas y principalmente en la superficie oclusal de los molariformes, que se atribuye a la variación intraespecífica que se presenta de forma natural en una población; esto a su vez su vez refleja la presencia de individuos de diferente edad y sexo entre los ejemplares estudiados. T. hicksi se encuentra en la Cuenca de Tecolotán asociado con Dinohippus mexicanus, Nannippus aztecus, Neohipparion eurystyle, Astrohippus stockii y Borophagus secundus, por lo que la edad de la fauna corresponde al Henfiliano tardío.
Agradecimientos
Quisiéramos expresar nuestro agradecimiento a Diana Solórzano Pérez, del Museo de Paleontología de Guadalajara "Federico Solórzano Barreto" y a Martelva Gómez Pineda, del Museo Regional de Guadalajara, por darnos acceso al material de rinocerontes que tienen bajo su resguardo. Ma. del Carmen Perrilliat, encargada de la Colección Nacional de Paleontología, amablemente dio acceso al material de rinocerontes proveniente de El Ocote, Guanajuato. Agradecemos también a Óscar J. Polaco, quien contribuyó con valiosos comentarios en la primera versión del manuscrito, a Óscar Carranza–Castañeda y Esperanza Cerdeño, quienes con sus atinadas sugerencias ayudaron a mejorar este trabajo. G. Carbot–Chanona agradece al proyecto FOMIX CHIS–2005–C03–020 y al Instituto de Historia Natural por financiar la investigación, y a Francisco J. Vega (Instituto de Geología, UNAM) por su valioso apoyo durante el proceso de arbitraje.
Referencias
Brunet, J., 1968, The Pliocene Rhinoceroses of Mexico (resumen), en Geologic Society of America Annual Meetings, Paper 12, Mexico City, 1–2.
Carbot– Chanona,G., 2008, The first record of Crocodylus (Crocodylomorpha, Crocodylidae) from the Latest Miocene of Chiapas, Mexico, en III Congreso Latinoamericano de Paleontología de Vertebrados, Neuquén, Argentina.
Carranza–Castañeda, O., 1989, Rinocerontes de la Fauna Local Rancho El Ocote, Mioceno tardío (Henfiliano tardío) del estado de Guanajuato: Revista del Instituto de Geología, Universidad Nacional Autónoma de México, 8(1), 88–99.
Carranza–Castañeda, O., 1992, Una nueva localidad del Henfiliano tardío en la Mesa Central de México: Revista del Instituto de Geología, Universidad Nacional Autónoma de México, 10(2), 179–196.
Carranza–Castañeda, O., 2006, Late Tertiary fossil localities in Central Mexico, between 19°–23°N", en Carranza–Castañeda, O., Lindsay E.H. (eds), Advances in late Tertiary vertebrate paleontology in Mexico and the Great American Biotic Interchange: Universidad Nacional Autónoma de México, Instituto de Geología y Centro de Geociencias, Publicación Especial, 4, 45–60.
Carranza–Castañeda, O., Ferrusquía–Villafranca, I., 1978, Nuevas investigaciones sobre la fauna Rancho El Ocote, Plioceno medio de Guanajuato, México; informe preliminar: Revista del Instituto de Geología, Universidad Nacional Autónoma de México, 2(2), 163–166.
Castillo–Cerón, J., Cabral–Perdomo, M.A., Carranza–Castañeda,O., 1996, Vertebrados fósiles del Estado de Hidalgo: Universidad Autónoma del Estado de Hidalgo, 127 p.
Cook,H. J., 1927, Anew rhinoceros of the genus Teleoceras from Colorado: Proceedings of the Colorado Museum of Natural History, 7, 1–2.
Cook, H.J., 1930, New rhinoceroses from the Pliocene of Colorado and Nebraska: Proceedings of the Colorado Museum of Natural History, 9,44–51.
Cope, E.D., 1884, The extinct mammalian of the Valley of Mexico: Proceedings of the American Philosophical Society, 22(7), 1–21.
Dalquest, W.W., Mooser, O., 1980. Late Hemphillian mammals of the Ocote local fauna, Guanajuato, Mexico: Texas Memorial Museum, The Pearce–Sellards Series, 32, 1–25.
Ferrusquía–Villafranca, I., 1990, Contribución al conocimiento de los mamíferos miocénicos de Chiapas, México: Revista de la Sociedad Mexicana de Paleontología, 2(2), 77–91.
Freudenberg, W., 1921, Geologie von México: Verlag von Geobrüder Borntraeger, Berlin, 1–32.
Freudenberg, W., 1922, Die säugetierfauna des Pliocäns und Postpliocäns von Mexiko. II Teil, Mastodonten und Elephanten: Geologie und Paläontologie, 14(3), 103–176.
Hatcher, J.B., 1894, Amedian horned rhinoceros from the Loup Fork beds of Nebraska: American Geologist, 13(3), 149–150.
Kowallis, B.J., Swisher, C.C., Carranza–Castañeda, O., Miller, W.E., Tingey, D.G., 1998, Fision–track and single–crystal 40Ar/39Ar laser–fusion ages from volcanic ash layers in fossil–bearing Pliocene sediments in Central Mexico: Revista Mexicana de Ciencias Geológicas, 15(2), 157–160.
Lance, J.F., 1950, Paleontología y estratigrafía del Plioceno de Yepómera, Estado de Chihuahua, parte I: Equidos, excepto Neohipparion: Boletín del Instituto de Geología, Universidad Nacional Autónoma de México, 5, 87 p.
Lindsay, E.H., 1984, Late Cenozoic mammals from northwestern Mexico: Journal of Vertebrate Paleontology, 4(2), 208–215.
Maldonado–Köerdell, M., 1954, Sobre un rinoceronte fósil de Sinaloa, México: Sociedad Mexicana de Historia Natural, 5(15), 141–146.
Matthew, W.D., 1932, A review of the rhinoceroses with a description of Aphelops material from the Pliocene of Texas: Bulletin of the Department of Geological Sciences, University of California Publications, 20, 411–480.
Mead, A.J., 2000, Sexual dimorphism and paleoecology in Teleoceras, a North American Miocene rhinoceros: Paleobiology, 26(4), 689–706.
Mihlbachler, M.C., 2005, Linking sexual dimophism and sociality in rhinoceroses: insights from Teleoceras proterum and Aphelops malacorhinus from the Late Miocene of Florida: Bulletin of Florida Museum Natural History, 45(4), 495–520.
Miller, W.E., Carranza–Castañeda, O., 1998, The Tecolotlán Basin, Central Mexico: Anew area for Late Cenozoic terrestrial vertebrates: Journal of Vertebrate Paleontology, Supplement, 18(3), 26.
Miller, W. E., Carranza–Castañeda, O., 2002, Importance of Mexico's late Tertiary mammalian faunas, en Montellano–Ballesteros, M., Arroyo– Cabrales, J. (eds.), Avances en los estudios paleomastozoológicos: México, D.F., Instituto Nacional de Antropología e Historia, 83–102.
Montellano–Ballesteros, M., 1997, New vertebrate locality of the late Hemphillian age in Teocaltiche, Jalisco, Mexico: Revista Mexicana de Ciencias Geológicas, 14(1), 84–90.
Osborn, H.F., 1898, A complete skeleton of Teleoceras fossiger. Notes upon the growth and sexual characters of this species: Bulletin of American Museum of Natural History, 10, 51–59.
Osborn, H.F., 1904, New Miocene rhinoceroses with revision of known species: Bulletin of American Museum of Natural History, 20(27), 307–326.
Prothero, D.R., 2005, The Evolution of North American Rhinoceroses: Cambridge University Press, 218 p.
Rodríguez–Huerta, M., Guzmán–Gutiérrez, R., 1993, Fauna de vertebrados fósiles del Mioceno tardío del área de Tecolotlán, Jalisco, México, en IV Congreso Nacional de Paleontología: Sociedad Mexicana de Paleontología, 50.
Tanner, L.G., 1975, Stratigraphic occurrences of Teleoceras, with a new Kimballian species from Nebraska: Bulletin of the University of Nebraska State Museum, 10, 23–33.
Manuscrito recibido: Febrero 27, 2009.
Manuscrito corregido recibido: Junio 23, 2009.
Manuscrito aceptado: Junio 28, 2009.