
|
Boletín de la Sociedad Geológica Mexicana Volumen 75, núm. 2, A260423, 2023 http://dx.doi.org/10.18268/BSGM2023v75n2a260423
|
 |
Biogeoquímica isotópica y desgaste dental en Mammuthus columbi: Análisis multiproxy y revisión para su reconstrucción paleodietética
Isotope biogeochemistry and tooth wear in Mammuthus columbi: Multiproxy analysis and review of its paleodiet reconstruction
Susana Rodríguez-Franco1,*, Víctor Adrián Pérez-Crespo 2, Florent Rivals 3,4,5
1 Posgrado en Ciencias Biológicas, Universidad Nacional Autónoma de México, Edificio A, 1° Piso, Circuito de Posgrados, Ciudad Universitaria, 04510, Coyoacán, CDMX, México.
2 Instituto de Geología, Universidad Nacional Autónoma de México, Ciudad Universitaria, Circuito de la Investigación Científica S/N, 04510, Coyoacán, CDMX, México.
3 ICREA, Pg. Lluís Companys 23, 08010 Barcelona, España.
4 Institut Català de Paleoecologia Humana i Evolució Social (IPHES-CERCA), Zona Educacional 4, Campus Sescelades URV (Edifici W3), 43007, Tarragona, España.
5 Universitat Rovira i Virgili, Departament d’Història i Història de l’Art, Avinguda de Catalunya 35, 43002, Tarragona, España.
* Autor para correspondencia:(S. Rodríguez- Franco) This email address is being protected from spambots. You need JavaScript enabled to view it.
Cómo citar este artículo:
Rodríguez-Franco, S., Pérez-Crespo, V.A., Rivals, F., 2023, Biogeoquímica isotópica y desgaste dental en Mammuthus columbi:Análisis multiproxy y revisión para su reconstrucción paleodietética: Boletín de la Sociedad Geológica Mexicana, 75 (2), A260423. http://dx.doi.org/10.18268/BSGM2023v75n2a260423
Manuscrito recibido: 14 de Febrero de 2023; Manuscrito corregido: 26 de Abril de 2023; Manuscrito aceptado: 30 de Abril de 2023.
RESUMEN
En los últimos años, los estudios paleodietéticos y paleoambientales han tenido un gran avance debido a nuevas técnicas, como el análisis del desgaste dental a nivel micro y macro y, el análisis de isotopos estables de diferentes elementos en los tejidos de los animales. Recientemente, la combinación de éstos proxies ha mejorado sustancialmente este tipo de estudios, sin embargo, estos análisis van de la mano con la biología y comportamiento animal, por lo que es fundamental primero comprender a las especies estudiadas. En el presente trabajo se describen los análisis comúnmente utilizados en los últimos años en el mamut columbino (Mammuthus columbi) y la relación que tiene la biología de esta especie con la correcta aplicación de estos análisis. Así mismo, las muestras y medidas tomadas de estos individuos fósiles deben tomarse con cuidado, ya que las peculiaridades en la ontogénesis dental, de comportamiento y metabolismo, son clave para la comprensión de los resultados obtenidos.
Palabras clave: reconstrucción dietética, mesodesgaste dental, microdesgaste dental, Isotopos estables, Pleistoceno final, Proboscidia.
ABSTRACT
In recent years, paleodietary and paleoenvironmental studies have made great progress due to new techniques, such as the analysis of dental wear at micro and macro level and the analysis of stable isotopes of different elements in animal tissues. Recently, the combination of these proxies has substantially improved these types of studies, however, these analyses go hand in hand with biology and the animal behavior, so it is essential to understand the species studied previously. This paper describes the analyses commonly used in recent years in the Columbian mammoth (Mammuthus columbi) and the relationship that the biology of this species has with the correct application of these analyses. Likewise, the samples and measurements taken from these fossil individuals must be taken with care, since the peculiarities in dental ontogenesis, behavior and metabolism are key to the comprehension of the results obtained.
Keywords: dietary reconstruction, dental mesowear, dental microwear, stable isotopes, Late Pleistocene, Proboscidia.
- Introducción
Las inferencias paleodietéticas han ayudado a conocer las fuentes de alimentación de las especies extintas y la manera en que se alimentaban para su supervivencia, la gama de recursos alimentarios disponibles para una especie es también una indicación de las características físicas y ecológicas de los hábitats a los que se adapta una especie, y así mismo, los cambios dietéticos individualmente o de comunidades enteras, pueden reflejar cambios ecológicos y evolutivos (Fox y Fisher, 2004).
Algunos megaherbívoros del Pleistoceno Tardío son considerados como “herbívoros clave”, debido a que superaban los 1000 kg de masa corporal, poseyendo una invulnerabilidad a la depredación (no humana) de los individuos adultos, alcanzando grandes densidades de saturación (Owen-Smith, 1987). La capacidad de estos megaherbívoros de consumir grandes cantidades de plantas, tiene consecuencias directas en la vegetación y en los suelos (Lundgren et al., 2021). La reducción y extinción de éstos megahebívoros durante el Pleistoceno tardío significó un importante cambio en la estructura de los paisajes y los ecosistemas del mundo, teniendo una importante participación en la regulación de las plantas leñosas (Bakker et al., 2016).
Esta importancia ecológica puede observarse actualmente en elefante africano (Loxodonta africana), los cuales tienen la capacidad de cambiar bosques densos o zonas de matorrales en pastizales, modificando en gran medida el hábitat donde viven (Owen-Smith, 1987). Por ello, son llamados “ingenieros del ecosistema”, su actividad biológica puede transformar el paisaje, modificando incluso el suelo, a causa del pisoteo continuo por sus rutas migratorias, dejando senderos a su paso, comúnmente llamados “senderos de elefantes”, y que pueden ser usados por estos últimos durante generaciones (Haynes, 2012). Así mismo, las preferencias dietéticas de los proboscidios están relacionadas directamente con la disponibilidad de los diferentes tipos de vegetación, y han demostrado poseer la capacidad de adaptarse a los cambios de vegetación en su hábitat (Cerling et al., 1999). Por lo que los proboscidios fósiles han sido de gran ayuda en el estudio de transiciones climáticas y de cambios de vegetación a través del tiempo (Cerling et al., 1999; Camidge et al., 2020).
El estudio del comportamiento y la dieta de las especies fósiles son clave para las reconstrucciones ambientales y ecológicas, y las cuales, anteriormente habían sido inferidas mediante la morfología de sus estructuras óseas, sin embargo, actualmente el análisis de diferentes proxies, tales como el desgaste dental y el análisis de isotopos estables de carbono y oxígeno en esmalte dental y, de carbono y nitrógeno en colágeno, han ayudado a efectuar mejores y más precisas inferencias (Metcalfe et al., 2013; Lister, 2014; Metcalfe, 2017; González-Guarda et al., 2018; Uno et al., 2020; Cherney et al., 2021; Wooller et al., 2021; Miller et al., 2022). En los últimos años, estas técnicas han demostrado ser eficaces para la comprensión de cómo se alimentaban los proboscidios y como cambió la disponibilidad de las plantas que consumían, así como de los cambios ambientales que ocurrieron en el pasado (Iacumin et al., 2010; Saarinen et al., 2015; Schwartz-Narbone et al., 2019; Camidge et al., 2020, Smith y DeSantis, 2020; Liu et al., 2022). Adicionalmente, los análisis isotópicos, especialmente los del oxígeno, no sólo son capaces de arrojar información sobre el tipo de alimentación y hábitat, sino que también han demostrado ser eficaces para las inferencias paleoclimáticas locales, haciendo posible el estudio de migraciones (Longinelli, 1984; Tüken et al., 2007; Uno et al.,2020b; Wooller et al., 2021). Por lo anterior, el estudio de los proboscidios puede ayudar a mejores interpretaciones ecológicas y ambientales, tanto en ecosistemas actuales como del pasado, sin embargo, para su correcta interpretación, es importante conocer las técnicas disponibles y sus alcances, así como su correcta aplicación. En el presente artículo se reúnen las técnicas más utilizadas actualmente para el análisis paleodietético de Mammuthus columbi en México, estos son: análisis de isotopos estables de carbono y oxígeno en esmalte dental, de carbono y nitrógeno en colágeno, y análisis de desgaste dental a nivel micro y macro. Así mismo, se presentan las peculiaridades biológicas de los proboscidios que podrían ayudar a entender mejor éstos análisis, así como las problemáticas que pudieran presentarse al estudiar estos grandes mamíferos.
- Mammuthus columbi
Los mamuts (familia Elephantidae, género Mammuthus) pertenecen al orden Proboscidea, al cual también forman parte los únicos dos géneros conocidos de elefantes modernos, Loxodonta y Elephas. En la actualidad estos grandes mamíferos se distribuyen únicamente en el continente africano y asiático respectivamente, sin embargo, durante el Pleistoceno Tardío, existían tres familias de proboscidios en América: Gomphotheriidae (gonfoterios), Mammutidae (mastodontes) y Elefantidae (mamuts), los cuales se extinguieron durante la transición Pleistoceno-Holoceno hace unos 10 a 11 mil años AP (antes del presente) (Arroyo-Cabrales et al., 2007).
El género Mammuthus se distribuyó desde Canadá, Estados Unidos de América, México, y parte de Centroamérica, no obstante, éstos grandes mamíferos no se distribuyeron más allá de Panamá (Arroyo-Cabrales et al., 2007). Los primeros mamuts en el continente americano aparecen hace aproximadamente 1.5 millones de años, estos probablemente evolucionaron desde un mamut euroasiático, el mamut de la estepa (Mammuthus trogontherii), el cual cruzó el estrecho de Bering y dio lugar primeramente, a Mammuthus columbi (Mamut colombino) y posteriormente a Mammuthus primigenius (Mamut lanudo) en Beringia, este último restringió su distribución al norte de América y Europa, mientras que M. columbi tuvo una amplia distribución en el continente Americano (Lister y Sher, 2015). Adicionalmente, en la isla de Santa Rosa, en el archipiélago de California, Norte América, vivió una especie de mamut de pequeñas dimensiones, llamado mamut pigmeo (Mammuthus exilis). Estos mamuts debieron haber evolucionado desde poblaciones de M. columbi que cruzaron a este sistema de islas cuando el nivel del mar era bajo, sufriendo un enanismo insular debido al aislamiento en estas pequeñas islas (Agenbroad, 2001, 2003; Muhs et al., 2015).
2.1. ELEFANTES ACTUALES Y EL MAMUT COLOMBINO
Los elefantes actuales (Elephas y Loxodonta) poseen un comportamiento social muy estructurado y complejo, los machos adultos son solitarios o viven en pequeñas manadas de elefantes solteros (Schulte, 2000), por otro lado, los elefantes hembras viven en unidades familiares llamadas matriarcales, compuestas principalmente por hembras adultas y crías, ésta unidad familiar está dirigida por la hembra más longeva del grupo (matriarca) y en la cual, la experiencia y edad juega un papel importante en el éxito reproductivo de todas las hembras de la unidad familiar (McComb et al., 2001), además de que las decisiones de la matriarca influyen totalmente en el comportamiento de esta unidad familiar (Moss, 2012).
Además de las diferencias de comportamiento que muestran ambos sexos, también las hay en los hábitos alimenticios, que incluso existen entre individuos juveniles y adultos. Algunos estudios indican que en L. africana, existe una marcada diferencia de alimentación entre hembras y machos adultos, siendo las hembras las que mantienen una dieta más selectiva y nutritiva, ingiriendo una cantidad mayor de plantas leñosas, contrario a esto, los machos presentan una alimentación mucho más generalista (Stokke, 1999). Por otro lado, Sukumar y Rames (1992) y Greyling (2004) indican que, tanto en los elefantes africanos como en asiáticos, los individuos adultos integran a su dieta una mayor cantidad de árboles y arbustos que los jóvenes, los cuales tienen una preferencia al consumo de pastos.
No solamente se han observado estas variaciones dietéticas, existen evidencias que apuntan a que los elefantes actuales modifican su dieta a lo largo del año y esta se haya relacionada con los cambios de vegetación que ocurren estacionalmente, así, en la temporada de lluvia existe un aumento en la integración de plantas C4 y en cambio, cuando la estación seca llega, comienzan a alimentarse mayoritariamente de plantas C3 (Wittemyer et al., 2009; Codron et al., 2011; Koirala et al., 2016; Uno et al., 2020).
Actualmente existen algunas evidencias que podrían apuntar a que los mamuts pudieron tener un comportamiento muy similar al de los elefantes actuales y, por lo tanto, también en su manera de alimentarse (Haynes, 1993; 2012). Por ejemplo, los “mammoth rub sites” han sido interpretados como probables lugares donde los mamíferos del Pleistoceno (entre estos, mamuts) rascaban o frotaban sus cuerpos. Uno de estos lugares, ubicado en la costa norte de California, Estados Unidos, específicamente en el condado de Sonoma, donde, por medio de un análisis de micrografías (Microscopia de fuerza atómica (AFM)), se comparó la superficie de las “mammoth rub sites” y las rocas que son utilizadas por elefantes actuales para frotar sus cuerpos, encontrandose grandes similitudes entre ambos (Parkman 2002; 2009).
Adicionalmente, Saunders (1999), basándose en los hallazgos del yacimiento de Hot Springs en Dakota del Sur, Estados Unidos, sitio donde se han encontrado más de 50 individuos pertenecientes a Mammuthus columbi y Mammuthus primigenius datados en 26 000 A.P. y, los cuales en su mayoría eran adultos machos, solitarios y que fallecieron en momentos diferentes, propone la similitud que existe entre la estructura social de M. columbi y de proboscidios actuales. Por otro lado, en el yacimiento de Dent, en The Waco Mammoth National Monument (WMNM) en el centro de Texas, Estado Unidos, se han encontrado 25 individuos de Mammuthus columbi que probablemente representan el único grupo social encontrado hasta la fecha, ya que podría estar representada una manada matriarcal que migraba hacía fuentes de agua durante una intensa sequía (Haynes et al., 1992; Wiest et al., 2016).
- Análisis isotópicos de oxígeno, nitrógeno y carbono en bioapatita de mamíferos
Usualmente, los tejidos mineralizados fósiles como los huesos y dientes, son los que se conservan mejor a través del tiempo, por ello, son comúnmente analizados para realizar estudios isotópicos de carbono, nitrógeno y oxígeno (Koch, 2007; Zazzo et al., 2012). Esto es debido a que presentan una importante resistencia a las alteraciones químicas y biológicas que ocurren postmortem (aproximadamente hasta 108 años); estos tejidos están compuestos de proteínas, lípidos, y principalmente de una forma altamente sustituida de hidroxiapatita (Ca10[PO4]6[OH]2) llamada bioapatita, la cual ha integrado a su estructura iones carbonato (CO32-), entre otros (LeGeros, 1991; Koch, 2007). El hueso y la dentina contienen una composición similar, aproximadamente un 70% de cristales de bioapatita y su resto pertenece a una matriz orgánica de colágeno proteico (Lowenstam y Weiner 1989; Simkiss y Wilbur, 1989; Hedges, 2003; Morales-Puente et al., 2012). Una gran diferencia con la estructura del esmalte, el cual sólo aproximadamente el 5% de su peso pertenece a materia orgánica (LeGeros, 1991).
3.1. CARBONO
En las plantas, pueden presentarse tres tipos de vías fotosintéticas, el Ciclo de Calvin (C3), la vía Hatch-Slack (C4) y CAM (Metabolismo Ácido Crasuláceo) y las cuales, se presentan con frecuencia en tipos de plantas específicas, por ejemplo, la mayoría de las gramíneas poseen una vía C4 y, en cambio, la mayoría de árboles y arbustos tienen una vía C3 (Stowe y Teeri, 1978; Medrano y Flexas, 2000; Morales-Puente et al., 2012). Así mismo, estas vías metabólicas pueden identificarse mediante el análisis de isotopos estables del carbono de los tejidos de las plantas, la vía C3 tendra valores de δ13C dentro de un intervalo de -22‰ a -30‰, por otro lado, la vía C4 se encontrará dentro del intervalo de -10‰ a -14‰ (Farquhar et al., 1989; Steuter et al., 1990; Cerling et al., 1997).
En el análisis isotópico de carbono es importante considerar diversos factores que pudieran hacer variar los valores de δ13C que exhiben las plantas, uno de ellos es el CO2 atmosférico, que es su fuente principal de carbono (Koch, 1998). Y en el cual, la composición isotópica del carbono ha cambiado a lo largo del tiempo, desde un valor de δ13C de -6.5 durante el Pleistoceno, hasta -8.4‰ en la actualidad, esto debido a la intensiva quema de combustibles fósiles (Tipple, 2010; Keeling et al., 2017).
Otros factores que deben tomarse en cuenta en este análisis, es la eficiencia con la que se fija carbono por medio de la fotosíntesis, las plantas C4 poseen una mejor eficiencia que las plantas C3 y además, una mejor adaptación a soportar otras situaciones de estrés (Edwards et al., 2010; Gowik y Westhoff, 2011; Way et al., 2014). La abundancia de plantas C4 se correlaciona con temperaturas altas y baja humedad (Stowe y Teeri, 1978; Morales-Puente et al., 2012). Por otro lado, los valores isotópicos en las plantas C3 son muy susceptibles a bajos niveles de CO2 atmosférico, luz, estrés hídrico y osmótico, niveles de nutrientes en el suelo, aridez y temperatura (Ehleringer y Monson, 1993; Koch, 1998). Se ha observado que la abundancia de plantas C3 incrementa en zonas con temperaturas por debajo de los 25°C (Medrano y Flexas, 2000).
Por lo anterior, la precipitación media anual también es un factor muy importante en la interpretación de los valores isotópicos obtenidos, las plantas C3 pueden mostrar un sesgo con valores correspondientes a plantas C4 y CAM en ecosistemas secos y desérticos, una precipitación menor a 500 mm/año, causaría que las plantas posean valores mayores a -25.5‰, así mismo, valores tan negativos como -31.5‰ indicarían un bosque denso de dosel cerrado con poca luz y abundantes precipitaciones (Kohn, 2010).
Por otro lado, la plantas que habitan en bosques densos tienden a mostrar valores más negativos que los de hábitats abiertos y de pastizales, esto se conoce como efecto dosel y es provocado por un reciclaje del carbono ya fraccionado a partir de la descomposición de la materia orgánica del suelo del bosque, por lo tanto, valores muy negativos de δ13C indican un efecto dosel más marcado y hábitats más cerrados (Vogel, 1978; Van der Merwe y Medina, 1991; Bonafini et al., 2013).
Cuando los mamíferos herbívoros se alimentan, incorporan a sus tejidos las formas isotópicas estables del carbono, incluyendo las formas más abundantes, 12C y 13C. Anteriormente, se consideraba que los carbonos que constituyen los carbonatos del esmalte dental, mostrarían valores de δ13C más positivos en un 14‰ que las plantas consumidas y, en el caso de la bioapatita ósea, 5‰ más positivos que su alimento (Cerling y Harris, 1999; Cerling et al., 2005). Sin embargo, recientemente se ha demostrado que este incremento en los valores de δ13C varía por la fisiología de cada animal y principalmente, de su tamaño corporal (Tejada-Lara et al., 2018). Por otro lado, en el caso del colágeno óseo, se ha documentado que organismos con una preferencia de alimentación a plantas C3 (árboles, arbustos y pastos de zonas frías) mostrarán valores de δ13C en un intervalo de -28‰ a -18‰, y de -7‰ a -11‰ para las muestras provenientes de hueso perteneciente a animales que se alimentaron mayoritariamente de plantas C4 (pastos y algunas plantas de zonas cálidas) (MacFadden y Cerling, 1996).
3.2. NITRÓGENO
Las dos formas isotópicas estables más abundantes y comunes del nitrógeno son 14N y 15N, y las cuales, son incorporadas por las plantas desde el suelo hasta sus tejidos, por lo que éstos últimos reflejaran las proporciones isotópicas del nitrógeno provenientes del suelo, sin embargo, pueden tener grandes variaciones en sus valores δ15N, que van desde -8‰ y hasta +18‰ en las plantas desérticas (Shearer et al. 1983; Schoeninger y DeNiro 1984; Kelly, 2000). Esto se debe principalmente a la gran variación de dichos valores en los suelos y adicionalmente, a algunas estrategias que poseen las plantas para obtener este elemento, como es la presencia de mecanismos de fijación del nitrógeno, por lo que las plantas fijadoras de nitrógeno exhiben evidentemente valores de δ15N más negativos que las plantas que no tienen este mecanismo (Schoeninger y DeNiro, 1984; Kelly, 2000).
Los herbívoros adquieren nitrógeno de las plantas consumidas y así mismo, los carnívoros lo adquieren de sus presas (DeNiro y Epstein, 1981). El nitrógeno se incorpora a la estructura de muchos de sus tejidos, uno de ellos es él colágeno, el cual forma parte de la matriz orgánica de los huesos y la dentina. El colágeno es extraído para el análisis isotópico del nitrógeno, ya que se ha identificado que existen cambios en las proporciones isotópicas respecto a la cantidad de proteínas de origen animal y vegetal que se incluyen la alimentación de un animal, contribuyendo así, al enriquecimiento isotópico del nitrógeno en el colágeno, actualmente conocido como factor de enriquecimiento trófico (TEF en inglés) (Hobson y Bairlein, 2003; Krajcarz et al., 2018). Esta señal isotópica ha sido muy útil en las reconstrucciones tróficas para especies fósiles, ya que en general, los valores de δ15N del colágeno de los mamíferos son más positivos que el alimento consumido en un 3‰ a 5‰ por cada paso hacia arriba de los niveles tróficos (DeNiro y Epstein, 1981; Bocherens y Drucker, 2003; Fox-Dobbs et al., 2008; Jaouen et al., 2022).
Las proporciones isotópicas del nitrógeno reflejadas en el colágeno de los mamíferos dependen directamente de los valores adquiridos por los herbívoros desde las plantas y, en el caso de los carnívoros y omnívoros, de las diferentes presas o fuentes de alimentación que posean (DeNiro y Epstein, 1981; Stephens et al., 2022). Ya que las plantas son el medio por el que comienza la incorporación del nitrógeno en la red trófica, es de gran importancia considerar las condiciones ambientales y ecológicas donde viven o vivieron los mamíferos, por ejemplo, el nitrógeno de las plantas que habitan lugares áridos normalmente son más positivos que los de lugares húmedos y, contrario a esto, los valores son más positivos en sitios costeros o salinos que, los valores de las plantas que habitan sitios tierra adentro o no salinos (Heaton, 1987). Así mismo, se ha documentado que los valores de δ15N en el colágeno de mamíferos que habitan sitios cálidos y áridos son más positivos que los que habitan ambientes fríos y húmedos (Ambrose, 1991). Por otro lado, el consumo preferencial de algunos tipos de plantas tiene una evidente influencia en los valores de δ15N del colágeno, ya que son comúnmente bajos en los mamíferos que se alimentan de plantas fijadoras de nitrógeno, como, por ejemplo, las leguminosas (Delwiche et al., 1979; Ambrose y DeNiro, 1986; Koch et al., 1994).
Adicionalmente, los valores de δ15N reflejados en el colágeno óseo también variaran por la cantidad de lluvia que se precipite anualmente en su hábitat, y es especialmente evidente en áreas que reciben menos de 400 mm de lluvia por año (Sealy et al., 1987); de modo que, la señal isotópica del nitrógeno en el colágeno es inversamente proporcional a la cantidad de agua que consuman. Así, un estrés hídrico y una deshidratación incrementaría la concentración de urea en el cuerpo del mamífero y, en consecuencia, habría un incremento en el valor de δ15N (Ambrose y DeNiro, 1986).
Así mismo, la fisiología de los organismos también es un importante factor a tomar en cuenta en el análisis de las proporciones isotópicas del nitrógeno, las especies que poseen adaptaciones fisiológicas para una disminución de la ingestión de agua y su almacenamiento, tienen valores más positivos que las especies dependientes de este líquido (Ambrose, 1991). Otro ejemplo de esto, son los mamíferos que poseen un periodo de hibernación anual, como es el caso de algunos úrsidos, los cuales tienen un eficiente mecanismo de reciclaje de urea durante este periodo, y el cual, provocaría un aumento en los valores de δ15N en el colágeno (Ramírez-Pedraza et al., 2019; Krylovich et al., 2020). Adicionalmente, la lactancia que ocurre durante el crecimiento de las crías también puede incrementar los valores de δ15N en el colágeno, por lo que es un elemento valioso para la identificación del destete en los mamíferos (Jenkins et al., 2001; Cherney et al., 2021).
3.3. OXÍGENO
Existen variaciones naturales en la abundancia de los diferentes isótopos de oxígeno en las diferentes fuentes de agua de la superficie de la Tierra, dichas variaciones son provocadas por diferentes factores ambientales y geográficos, tales como: salinidad, altitud, latitud, longitud, precipitación, estacionalidad, evaporación y condensación y, temperatura (Epstein et al., 1951; Epstein y Mayeda, 1953; Dansgaard, 1964; Rozanski et al., 1993). Los valores isotópicos de oxígeno en el agua pueden ser tan específicos en cada sitio que es posible de crear mapas de datos con los valores de δ18O globales (Bowen et al., 2005).
El oxígeno se incorpora al hueso y esmalte de los mamíferos, mayoritariamente por medio del agua que es ingerida y consumida en los alimentos (Koch, 1998). El oxígeno adquirido es incorporado como carbonatos (CO32-) y fosfatos (PO43-) en la bioapatita (Koch, 2007; Morales et al., 2012). Por lo que, las proporciones isotópicas del oxígeno del agua consumida serán reflejejadas en la bioapatita, proporcionando información importante sobre el hábitat y el clima en el que vivieron los individuos (Kohn et al., 1996).
Un solo diente contiene información desde que comienza a formarse la pieza dental, hasta que emerge y detiene su mineralización (Higgins, 2018). Por lo anterior, el análisis de las proporciones isotópicas del oxígeno en la bioapatita ha sido muy útil para inferir los cambios ambientales y climáticos que ocurrieron a través del tiempo (Royer et al., 2013; Royer et al., 2014; Fernández-Garcia et al., 2022; Uno y Bibi, 2022
Por otro lado, con dicho análisis es posible identificar cambios en las fuentes de agua de los animales y, lo cual, podrían indicar movimientos migratorios (Bowen et al., 2005; Britton et al., 2009; Wooller et al., 2021; Miller et al., 2022). Así mismo, los valores de δ18O, son sensibles a los cambios estacionales, siendo especialmente útiles para la identificación de la estación en la que ocurrió la mineralizaron de los carbonatos en la bioapatita, esto es especialmente útil en el análisis del esmalte dental (Dutton et al., 2005).
3.4. FORMACIÓN DE LAS PIEZAS DENTALES DE LOS MAMUTS Y CONSIDERACIONES EN EL ANÁLISIS ISOTÓPICO DE C Y O EN EL ESMALTE
El desarrollo dental de los mamuts fue muy similar a la de los elefantes actuales, a lo largo de su vida presentaban seis diferentes molariformes en cada cuadrante de su cavidad bucal y de estos, sólo podían estar presentes uno o dos de en cada cuadrante de la mandíbula y maxilar al mismo tiempo (Laws, 1996). Estas piezas dentales avanzan en dirección anterior conforme surge el siguiente molariforme desde la parte posterior, por lo que todos los molariformes terminaban por caer por la parte anterior, normalmente con un marcado grado de desgaste oclusal. El orden del surgimiento, así como de la caída de los molares es el siguiente: M1→M2→M3→M4→M5→M6 (Laws, 1996). El surgimiento, el avance posterior-anterior y la muda de cada uno de los molares ha sido bien descrito y estudiado, por lo que se han establecido patrones de erupción, progresión dental y porcentaje de desgaste oclusal para la estimación de edades en Edad Elefante Africano (EEA) (Laws, 1966; Haynes, 1991).
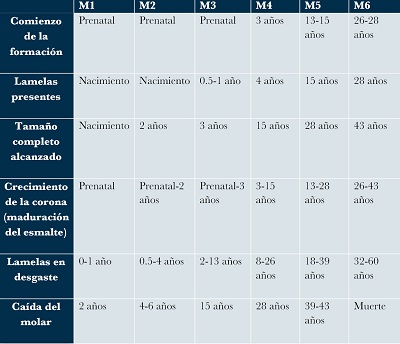
Además de lo anterior, los proboscidios (incluyendo mamuts) también presentan una formación especial de sus piezas dentales, éstos se desarrollan de una manera secuencial progresiva (de M1 a M6), primero, se forma la dentina, luego las placas de esmalte y por último el cemento sobre las coronas, esta formación empieza en la parte más anterior del molar y progresivamente se forma en dirección a la parte posterior. Las raíces se desarrollan después de que se ha formado el esmalte en todas las lamelas y, ésta secuencia anterior-posterior también se observa en las raíces al momento de la progresión del molar a la zona anterior de la mandíbula, las raíces anteriores se reabsorben mientras que las posteriores siguen siendo funcionales (Maschenko, 2002). La única excepción a esta formación secuencial es el molar M1, en el cual se forman simultáneamente todas las placas de esmalte, así como su corona, acompañadas por la formación del cemento el cual comienza su formación simultáneamente en la superficie de la corona y en la base de la raíz (Maschenko, 2002). Metcalfe et al. (2010), mencionan que los grandes molariformes pueden formarse durante años o incluso décadas y en consecuencia, la parte anterior puede estar en desgaste mientras que la anterior está aún en desarrollo, por lo que estos autores estimaron el tiempo que tarda en formarse completamente cada uno de los seis molares en los mamuts, basándose en la conocida propuesta de Laws (1996) para la estimación de la edad de los elefantes africanos (Loxodonta africana) (Tabla 1).
| Tabla 1. Edades estimadas en proceso de formación de los molares (Metcalfe et al., 2010). |
 |
Es importante indicar que la manera en que crece el esmalte en los proboscidios puede generar problemáticas y dudas en estudios paleodietéticos, paleoecológicos y paleoclimaticos, donde se analicen muestras del esmalte dental para el análisis isotópico del carbono y oxígeno. El proceso de formación y surgimiento de los dientes en los proboscidios posee una secuencia y tiempos específicos a lo largo de la estructura de cada molariforme, por lo que podría variar la información dietética entre los diferentes molares y a lo largo de cada uno de ellos. Metcalfe y Longstaffe (2017), estimaron que la tasa de crecimiento del espesor del esmalte es de entre 0.8 a 1.5 milímetros al año, así mismo, se estimó la tasa de crecimiento en altura de 13 a 14 mm/año. El esmalte comienza a formarse en la unión esmalte-dentina en la parte oclusal del diente y el crecimiento progresa en grosor hacia el exterior del diente (dirección contraria a la dentina) y en altura, en dirección a la base del diente, de tal forma que lo primero en formarse es el esmalte oclusal de cada diente (Hillson, 2005).
Por todo lo anterior, el muestreo “en bruto” del esmalte dental que comprenda todo el espesor de esta estructura, comprende al menos varios milímetros (Bulk samples, en inglés) y, la señal isotópica de estas muestras podría representar como mínimo varios meses y hasta varios años de formación de esmalte según sea el grosor de la muestra (Ferretti, 2003; Metcalfe et al., 2010). En consecuencia, si la muestra es tomada de la superficie oclusal del esmalte, la información contenida en ella será anterior a la información que contendrá una parte cercana a la raíz.
Por otro lado, los incisivos de los proboscidios, comúnmente llamadas “defensas” o colmillos”, se mudan una vez en su vida, la corona del primer par de incisivos termina de formarse a los 16 meses de formación y su raíz después de tres meses de su nacimiento, en cambio, los incisivos permanentes comienzan a formarse a los 17 meses de edad y no detienen su crecimiento durante toda la vida del animal (Raubenheimer, 2000; Raubenheimer et al., 1995)
Los incisivos presentan un crecimiento anual en su estructura en forma de láminas de dentina, éstas laminas pueden observarse como anillos de crecimiento anual a través de un corte transversal del incisivo, existen variaciones en el crecimiento de cada anillo, de tal manera que, puede identificarse un crecimiento más lento de la dentina durante la estación invernal, esto es, especialmente útil para la identificación de estacionalidades, cambios climáticos, migraciones y cambios de alimentación que pudieran ocurrir durante un año de vida (Koch et al., 1989; Rountrey et al., 2012). Por ejemplo, recientemente el análisis de isotopos de oxígeno y estroncio en un colmillo completo de M. primigenius evidenció los movimientos geográficos durante sus diferentes etapas de la vida (Wooller et al., 2021).
- Mesodesgaste dental
El análisis de mesodesgaste dental es un método relativamente reciente, que resulta bastante accesible en costos y que puede ser aplicado sin que los restos fósiles sufran daños mayores o destrucción completa de la pieza dental. Este análisis toma en cuenta el cambio en el desgaste total de la superficie oclusal de pieza dental que ocurre durante su uso, desde que emerge hasta que el diente cae o hasta que el individuo muere (Ackermans, 2020). Sin embargo, la información obtenida de este desgaste resulta poco específica para cortos periodos de tiempo (días o semanas) (Fortelius y Solounias, 2000; Rivals et al., 2013; Saarinen et al., 2015). A diferencia del desgaste que ocurre a nivel microscopico, donde se puede identificar el tipo de alimento que consumió el animal antes de morir o de haber perdido la pieza dental (Grine, 1986). Por lo tanto, el mesodesgaste permite hacer inferencias sobre la dieta de herbívoros fósiles a lo largo de su vida (Fortelius y Solounias, 2000) y, consiste en la observación, análisis y el grado en el que se desarrollan las facetas en el diente y como se relaciona este desgaste con la dieta consumida (Green y Croft, 2018).
El mesodesgaste original de Fortelius y Solounias (2000), se basó en la examinación y clasificación de los ápices de las cúspides de los molares presentes en los artiodactylos y perissodactylos, específicamente los bordes bucales de los paraconos y metaconos, caracterizando el relieve y la forma de las cuspides. El relieve de las cúspides se refiere a la diferencia de relativa de altura entre las puntas de las cúspides y los valles entre las cúspides, es decir, la altura de las cúspides en la vista lateral, clasificando esta altura en alto o bajo. Por otro lado, la forma de la cúspide se refiere al vértice de la cúspide descrito como afilado, redondeado o romo, en orden decreciente de desarrollo de facetas. Con esta clasificación de cúspides, pueden clasificarse a los hervíboros en cuatro grupos principales: pacedores, consumidores mixtos con un dominio pacedor, consumidores mixtos dominados por ramoneo y finalmente, ramoneadores.
El desgaste dental puede ocurrir de dos formas diferentes, por medio de abrasión o por atrición (Fortelius, 1985; Fortelius y Solounias, 2000; Green y Croft, 2018). La abrasión es el resultado del desgaste causado por alimentos abrasivos que desgastan el diente, existe evidencia sobre el importante desgaste dental que provoca una dieta basada en pastos, los cuales, contienen una gran cantidad de silicatos en su estructura celular llamados fitolitos, que aunque la dureza de estas estructuras son menores al del esmalte, contribuyen de manera importante al desgaste dental (Winkler et al., 2019); adicionalmente, una alimentación cerca del suelo podría contener sedimentos abrasivos como polvo y arena que podrían contribuir a un mayor desgaste (Baker et al., 1959; Janis y Fortelius, 1988; Kaiser et al., 2013, Ackermans, 2020). En un desgaste por abrasión, las cúspides se desgastan en una mayor proporción, siendo comúnmente bajas y romas. Por otro lado, la atrición, es consecuencia de un desgaste principalmente creado por el contacto entre dientes (desgaste diente-diente) y no por el alimento consumido, en consecuencia, las cúspides presentan un relieve oclusal alto y puntiagudo (Sanson, 2006). El desgaste por medio de atrición ocurre principalmente en dietas ramoneadoras (árboles, arbustos y frutos) (Fortelius y Solounias, 2000).
A pesar de que el mesodesgaste es un método bastante accesible, que facilita el análisis de un gran número de individuos, el desgaste parece no ser estable a lo largo de la vida de los individuos y debe considerarse la edad ontogénica de los individuos estudiados, ya que se requiere que los dientes muestren facetas de desgaste bien definidas y sin daños (Green y Croft, 2018); en consecuencia, es importante excluir del análisis dientes demasiado desgastados y también los que se encuentran en una etapa de desgaste muy temprana (Fortelius y Solounias, 2000; Rivals et al., 2007). Aunado a esto, las tasas de mortalidad específicas de la especie también podrían tener una clara influencia en la señal de mesodesgaste, y adicionalmente, también es importante tener en cuenta si la muerte fue catastrófica o de forma natural (Rivals et al., 2007). Además, el esmalte de la superficie oclusal debe presentar un pulido suave y sin daños aparentes, lo que indicaría una buena conservación del esmalte (Green y Croft, 2018).
En el análisis de poblaciones de especies extintas y restos fósiles, muchas veces existe una gran problemática respecto a los pocos individuos disponibles para analizar. Green y Croft (2018) sugieren el análisis de al menos 10 individuos para poder obtener adecuadamente una variación en una población.
4.1. MODIFICACIONES Y AVANCES EN LA TÉCNICA DE MESODESGASTE
La facilidad, accesibilidad y rapidez del análisis de mesodesgaste dental ha posibilitado la modificación del método original de Fortelius y Solounias (2000). Kaiser et al. (2000) y Kaiser y Solounias (2003) fueron los primeros en extender las observaciones a más de una cúspide por individuo e involucrar a más de un observador en las mismas; a este método se le ha denominado como “mesodesgaste I”. Posteriormente Mihlbachler y Solounias (2006) propusieron un sistema de puntuación de cuatro estados categóricos, que culminó en el desarrollo de una escala de clasificación de siete estados, del 0 al 6, y que, mediante una comparación con esta escala, se combinaba la altura de la cúspide con el desgaste del ápice en una en una escala única, a este método se le llamo “mesodesgaste II” (Mihlbachler et al., 2011; Solounias et al., 2014). Más recientemente, la propuesta de Solounias et al. (2014) y Danowitz et al. (2016), en la cual, a través de un microscopio estereoscopio, se analiza por separado las superficies mesial y distal de la segunda banda de esmalte (más lingual), así como la unión entre las dos facetas, calificando las bandas de esmalte como navegadores cuando se caracterizan por ser “flat” y “planar”, en los pacedores si contiene gubias y sub-facetas que eventualmente se vuelven redondas. Mesodesgaste III califica estas formas con una escala de 1 a 4, indicando el número 1 un navegador típico y el número 4 un pacedor típico. Por último, Jiménez-Manchón et al. (2022) propusieron una modificación del “mesodesgaste II”, llamada “Mesodesgaste Dental Cuantitativo” en la cual se hacen mediciones del ángulo de desgaste y la altura de las cúspides a partir de fotografías, mejorando así, la categorización de los individuos de una manera más confiable en los siete estados categóricos propuestos en mesodesgaste II, además, este análisis abre la posibilidad de encontrar variantes alimentarias fuera de estas categorías.
Además de estas principales modificaciones del mesodesgaste original, hay variaciones de cada de éstas (Ackermans, 2020), por ejemplo, se han propuesto modificaciones para su aplicación en mamíferos de tamaño pequeño y mediano, como los lagomorfos (Fraser y Theodor, 2011), lepóridos y murinos (Ulbricht et al., 2015), campañoles (Kropacheva et al., 2017), marsupiales (Butler et al., 2014) y xenartros (Saarinen y Karme, 2017). También, se han propuesto análisis en grandes mamíferos, como los rinocerontes (Taylor et al., 2013) y proboscidios (Saarinen et al., 2015). Cada una de estas modificaciones ha demostrado ser efectiva para la investigación de la dieta en diferentes tipos y tamaños de dientes, así como en diversos mecanismos de masticación.
4.2. ANÁLISIS DE MESODESGASTE EN PROBOSCIDIOS
La metodología para el análisis de mesodesgaste dental en molares de proboscidios fue propuesta Saarinen et al. (2015), la cual es una modificación del “mesodesgaste II” para el análisis del mesodesgaste dental en los grandes molares que presentan los proboscidios. Debido al gran tamaño de las piezas dentales, además de la progresión dental que los proboscidios presentan, deben elegirse las tres lamelas centrales del diente y en caso de que el número de lamelas sea par, deberían elegirse una lamela anterior al centro del diente y dos posteriores del centro del mismo, esto con el objetivo de evitar errores de muestreo debido al desgaste excesivo de la superficie oclusal o al poco desgaste que presentan las lamelas posteriores debido a su poca progresión en la mandíbula.
A diferencia de otras técnicas de mesodesgaste, la manera de evaluar el desgaste de la superficie oclusal en los molares de los proboscidios es una aplicación del “mesodesgaste dental cuantitativo”, ya que, en vez de realizar comparaciones directas con categorías establecidas, se mide el ángulo de desgaste que ocurre desde la parte inferior de los valles dentales hasta la parte superior de las láminas de esmalte, esto puede ser medido por medio de un medidor de ángulos digital o un medidor de contornos convencional (Saarinen et al., 2015; Jiménez-Manchón et al., 2022) (Figura 1). Para un menor margen de error, los ángulos deben medirse en las tres lamelas centrales del diente y promediarse, para así obtener un ángulo general que represente al mesodesgaste total del molar. En el caso de la obtención de los ángulos por medio de un medidor de contornos, pueden medirse a través de una fotografía digital con ayuda de un programa de computadora con la opción de herramientas de medición de ángulos, por ejemplo, el programa de libre uso ImageJ versión 1.53 (2022).
 |
| Figura 1. Replicación de la superficie oclusal mediante un medidor de contornos convencional. |
Dado que los proboscidios presentan una alimentación bastante variada de tipos de plantas, con ayuda de esta propuesta, es posible diferenciar diferentes variabilidades de preferencias dietéticas de pastos o plantas leñosas. Así, ángulos >130° indicaría una alimentación estricta de plantas C4; de 130° a 124° pacedor pero con una mínima ingesta de un componente arbustivo; para los animales de dieta mixta (C3/C4), el intervalo va de 124° a 113°; aquellos que sean ramoneadores pero con un consumo mínimo de herbáceas, mostraran ángulos de 113° a 106° y, finalmente los consumidores estrictos de plantas C3 tendrán ángulos <106° (Saarinen et al., 2015).
4.2.1. Peculiaridades dentales
Una peculiaridad de los molares que poseen los proboscidios, es que a diferencia de otros grupos de mamíferos, donde el método de mesodesgaste se excluye a los inviduos con edades tempranas, debido al poco uso y desgaste de las piezas dentales, Cammidge et al. (2020) sugiere que en proboscidios no existe ninguna problemática al analizar el mesodesgaste de los individuos juveniles (M1 a M3) y adultos (M4 a M6).
Por otro lado, se ha observado que muchas veces los molares de los proboscidios pueden presentar malformaciones peculiares (Krzemińska, 2008; Grigoriev et al., 2014; Grigoriev et al., 2017) debido a un retraso en la ontogenia dental que caracteriza a la familia Elephantidae (Roth, 1989). Por lo que el desarrollo y progresión dental hacia la parte anterior de la mandíbula o maxilar es lenta, en consecuencia, muchas veces puede existir una malformación física de toda la pieza dental, provocando que los molares de los proboscidios sean más variables en longitud y anchura que otros mamíferos (Roth, 1992). Las fuerzas mecánicas de masticación, erupción y progresión que actúan en molares que están en desarrollo, pueden provocar una deformación importante de la superficie oclusal de los dientes de uno o ambos lados de la mandíbula o maxilar, estas malformaciones pueden provocar una mala oclusión, desalineamiento de los molares, y deformidades en la superficie oclusal (Roth, 1989).
A continuación (Figuras 2 a 5) se muestran molares y mandíbulas de Mammuthus columbi que se hallaron en la Cuenca de México, y en donde se ha podido identificar graves afectaciones de la superficie oclusal, consecuencia de malformaciones en las bandas de esmalte. El material fósil está albergado en la Colección Paleontológica del Laboratorio de Arqueozoología “M en C. Ticul Álvarez Solórzano” de la Subdirección de Laboratorios y Apoyos Académicos del INAH (Instituto Nacional de Antropología e Historia) (Figura 2 y 3), y en el Museo de la Casa de Cultura, Víctor Urbán Velasco (Figura 4 y 5), ambos en México.
 |
| Figura 2. Ejemplar con número de catálogo DP1948, proveniente de la Colección Paleontológica del Laboratorio de Arqueozoología “M en C. Ticul Álvarez Solórzano”. Presenta malformación en las primeras lamelas del molar, afectando de manera importante la forma y dirección de las láminas de esmalte. |
 |
| Figura 3. Ejemplar con número de catálogo 1825, de la Colección Paleontológica del Laboratorio de Arqueozoología “M en C. Ticul Álvarez Solórzano”. Presenta malformación de la superficie oclusal, con una pronunciada desviación del molar. |
Aunque hasta el momento no se ha comprobado si estas malformaciones podrían afectar el análisis de mesodesgaste, muchas veces los molares presentan una grave deformación y torsión de la superficie oclusal y podrían provocar diferencias en el desgaste de uno o ambos lados de la mandubula. Haynes y Klimowicz (2015) observaron desgaste extremo en los molares con malformaciones dentales, comparado a los que no presentaban malformaciones en la misma mandíbula. Adicionalmente, Grigoriev et al. (2017) encontraron un mayor desgaste en molares donde existe una importante torsión de las piezas dentales superiores, afectando incluso, las raíces. Por lo anterior, deben tomarse en cuenta estos cambios en la estructura del esmalte, y es recomendable no hacer inferencias dietéticas por medio de mesodesgaste en molares con malformaciones importantes de las láminas de esmalte.
 |
| Figura 4. Ejemplar número 470, albergado en el Museo de la Casa de Cultura, Víctor Urbán Velasco. Mandíbula con malformación en el molar derecho, afectando principalmente la forma de las lamelas y la profundidad de la superficie oclusal. |
 |
| Figura 5. Ejemplar número Tultepec I, albergado en el Museo de la Casa de Cultura, Víctor Urbán Velasco. Mandíbula con malformación en los molares izquierdos y derechos, afectando la dirección de su progresión dental. |
- Microdesgaste dental
A diferencia del mesodesgaste, esta técnica examina el desgaste que ocurre en el esmalte a nivel microscopico, este micro-desgaste es causado por el alimento al ser masticado y tiene el potencial de mostrar información sobre los últimos alimentos consumidos por el animal en días, horas e incluso, minutos antes de morir (Grine, 1986; Teaford y Oyen, 1989; Hoffman et al., 2015). Adicionalmente, se ha documentado que en algunos casos pueden observarse alteraciones alimentarias diarias, estacionales o regionales (Solounias y Semprebon, 2002; Semprebon y Rivals, 2010). La observación de las marcas (o cicatrices) en el esmalte de la superficie oclusal para la evaluación de las características dietéticas de los ungulados vivos y extintos, puede observarse mediante un microscopio de bajo aumento (35x); estas cicatrices pueden ser identificadas como gubias, fosas de diferente tamaño (pequeñas y grandes) y, lo mismo para los rayones (estrechos y anchos), y textura de los rayones (Teaford y Walker,1984; Solounias y Semprebon, 2002). Este micro-desgaste se correlaciona directamente con el tipo de alimento consumido, ya sea un alimento abrasivo o un alimento duro, por lo que puede diferenciarse un animal pacedor de un ramoneador (Walker et al., 1978). Solounias y Semprebon (2002) documentaron una base de datos con el análisis de microdesgaste dental de 50 especies diferentes de ungulados con dietas distintas y con diferentes patrones de desgaste, y con la cual, se facilitan comparaciones dietéticas entre ramoneadores, pacedores y consumidores mixtos.
Se ha documentado que el esmalte de herbívoros ramoneadores posee más fosas a causa de alimentos duros, muchas veces causados por ramas y cortezas (Semprebon et al., 2011; Rivals et al., 2019); en cambio, los herbívoros pacedores poseen más rayones en su esmalte dental a causa de la tierra y el pasto consumido, por otro lado, el esmalte de los organismos con una alimentación mixta tendrá patrones intermedios de fosas y rasguños (Solounias y Semprebon, 2002; Mainland, 2003; Asevedo et al., 2012).
La técnica de microdesgaste comenzó con las investigaciones de Mills (1955) y de Butler (1952), quienes investigaban la manera en que ocurría el movimiento oclusal de la mandíbula, mediante la observación de la superficie oclusal y desarrollo de las cúspides de los dientes. Posteriormente, los avances en la observación de la superficie del esmalte hallaron patrones de desgaste microscópicos (Rensberger, 1978), que proporcionaba información valiosa sobre las dietas de los animales y del hábitat donde vivían, por lo que el interés por descifrar mejor lo que significaban estas marcas contribuyó al avance de la observación por medio del microscopio electrónico de barrido (SEM, Scanning Electron Microscope). Además de esto, se evidenció la necesidad de una estandarización de la manera en que se observaban y documentaban los patrones de desgaste (Walker, 1978; Gordon, 1982; Teaford y Walker, 1984; Strait, 1993).
A pesar de que el análisis de microdesgaste dental mediante la microscopia óptica de luz tiene una relativa facilidad de uso a bajo costo, el método presentado inicialmente resultó ser lento, propenso al error por medición del observador y con una baja capacidad de repetibilidad, debido a la baja magnificación de imágenes bidimensionales que ofrecía el microscopio óptico, incluso, electrónicos de barrido. Por otro lado, éste último, resulto bastante costoso y poco viable para el estudio de un gran número de muestras (Solounias y Semprebon, 2002). En consecuencia, Solounias y Semprebon, (2002) desarrollaron una manera efectiva de la observación del microdesgaste mediante un estereomicroscopio, donde se mejoraba significativamente la estandarización de la observación de las marcas en el esmalte, por lo que resulto un método efectivo y de bajo costo, viable para un gran número de muestras y accesible. Partiendo de este método, con el tiempo se ha mejorado y se han propuesto soluciones respecto a los posibles errores de observación y capacidad de repetibilidad.
La observación del microdesgaste mediante la fotografía de alto rango dinámico (HDRI: High Dynamic Range Imaging) ha sido una de las soluciones para disminuir en gran medida el error del observador, la HDRI combina múltiples fotografías con exposiciones diferentes en una sola imagen, reduciendo las sombras y los reflejos no deseados, aumentando los detalles y, mejorando la visualización de las características del microdesgaste (Theodor y Furr, 2009; Fraser et al., 2009).
Por otro lado, también se han mejorado las observaciones mediante mediciones tridimensionales tomadas con microscopia confocal de luz blanca (white-light confocal microscopy, en inglés) junto con el análisis fractal sensible a la escala (Scale-sensitive fractal analysis), el cual está disponible como un software sensible a las texturas de las superficies analizadas (por ejemplo, http://www.surfract.com/). Esta metodología es la más utilizada actualmente, debido a que usa parámetros estandarizados para la caracterización de las cicatrices en el esmalte, por lo que ofrece caracterizaciones cuantitativas y repetibles de las superficies tridimensionales (Grine et al., 2002; Scott et al., 2005, 2006). Sin embargo, no son libres de error, los diferentes dispositivos utilizados para la captura de escaneos tridimensionales pueden producir algunas variaciones, incluso midiéndose la misma superficie (Winkler y Kubo, 2022).
Con el análisis mejorado de las texturas en la superficie del esmalte dental en tres dimensiones la técnica de microdesgaste no se limita sólo a herbívoros, si no que puede ampliarse a omnívoros y carnívoros, e incluso dentro de estos grupos identificar consumo de frutas, de huesos y/o de carne (p. ej., DeSantis, 2016; Pappa et al., 2019; Rivals et al., 2022).
5.1. MICRODESGASTE DENTAL EN MAMMUTHUS COLUMBI
El análisis de microdesgaste ha aportado de manera importante al comportamiento generalista de los proboscidios, revelando una dieta generalista flexible (Haiduc et al., 2018). Se ha estimado que la composición dietética reflejada por el microdestaste, en realidad exhibe las condiciones ambientales locales o regionales y no necesariamente la dieta “Optima adaptativa” (Zhang et al., 2017). Algo similar a lo que ocurre con los elefantes actuales, en los cuales el hábitat juega un papel importante en la alimentación de estos (Wittemyer et al., 2009; Codron et al., 2011; Haynes, 2012; Koirala et al., 2016; Uno et al., 2020). En consecuencia, estos mamíferos resultan bastante útiles para evaluar cambios ambientales, incluso, mostrando variabilidad entre poblaciones, claramente relacionada con los recursos disponibles y/o con limitaciones de competencia por los recursos con otros herbívoros (Rivals et al., 2019).
Un ejemplo de esta variabilidad alimenticia, es el estudio de Smith y DeSantis, (2018), en el que mediante el análisis de microdesgaste dental en mamuts colombinos y mastodontes (Mammut americanum) de Estados Unidos, han demostrado que pudieron ser similares en cuanto a las propiedades texturales de sus alimentos, siendo éstas dos especies tradicionalmente clasificadas como dietéticamente diferentes. Específicamente, los estudios recientes de microdesgaste en Mammuthus columbi han demostrado su amplia flexibilidad alimenticia, con una dieta tanto de ramoneo, como de pastoreo, pero, sin embargo, con una gran cantidad de fosas intensas comparado con otras especies de mamuts, lo que indica una ocupación de un hábitat más árido que otras especies (Rivals et al., 2019).
Para cualquier análisis de microdesgaste dental, comúnmente se fabrica una réplica de la superficie oclusal del diente para facilitar el traslado, disponibilidad y la observación de la superficie oclusal sin la necesidad de poseer físicamente el fósil original. En los molares de los proboscidios la creación de estas replicas son una muy buena opción, debido a que su peso no facilitan la observación en la mayoría de los microscopios. La réplica se fabrica comúnmente mediante el uso de impresión dental polivinilsiloxano de alta definición para la creación de la carcasa y resina epóxica para una réplica con una propiedad translúcida (Figura 6) que facilita la observación de las marcas en el esmalte (Solounias y Semprebon, 2002). La manera en que se realiza este procedimiento fue presentada de manera explícita por Martínez-Polanco et al. (2021).
 |
| Figura 6. Réplica de la superficie oclusal de un molar de Mammuthus columbi, fabricada de resina epóxica. |
Ya que los molares de los proboscídeos son de gran tamaño, el desgaste dental podría no ser representativo en una zona muy anterior o una zona muy posterior del molar, por lo que preferentemente debería elegirse las bandas de esmalte (lamelas) que se encuentran en la parte más central de la superficie oclusal (Solounias y Semprebon, 2002; Todd et al., 2007; Cammidge et al., 2020). Todd et al. (2007) y Rivals et al. (2012) recomiendan que el análisis de microdesgaste dental debe realizarse de esta manera debido a que el patrón de microdesgaste es más estable en las placas centrales que en la parte anterior, posterior, bucal y lingual. Adicionalmente, Cammidge et al. (2020) recomienda que para el análisis de microdesgaste dental en proboscideos fósiles (Mammut americanum y Mammuthus), si no es posible tomar el molde de los lofos de la zona media, pueden elegirse bandas alternas que se aproximen a éstas, prefiriéndose las bandas anteriores, donde el desgaste podría ser más aparente. Es importante asegurarse de fabricar una réplica del esmalte bien conservado y sin daño tafonómico aparente, aun así, podría no ser evidente la alteración del esmalte hasta el momento en el que esta siendo observado al microscopio, a continuación, se muestran dos ejemplos de un esmalte alterado y un esmalte bien conservado (Figura 7).
 |
| Figura 7. Derecha: microfotografía de esmalte bien conservado, se observa una superficie con aspecto suave y pueden identificarse fácilmente las cicatrices producidas por el alimento. Izquierda: microfotografía de esmalte mal conservado, posee una superficie estriada de manera regular, probablemente debido a procesos tafonómicos. |
El esmalte bien conservado siempre tendrá un aspecto “suave” cuando se mira al microscopio, por otro lado, muchas veces el esmalte mal conservado tendrá estriaciones muy pronunciadas y profundas, muchas veces con una superficie de apariencia rugosa o quebrada. Además, en un esmalte alterado las cicatrices no siguen la dirección de oclusión-masticación de la especie estudiada, en el caso de los proboscidios, debido a una masticación en dirección posterior-anterior (atrás-adelante), la mayoría de los rayones no deberían de tener una dirección izquierda-derecha. En la Figura 7, en la fotografía de la izquierda, puede identificarse fácilmente que la superficie del esmalte está totalmente afectada por algún elemento diferente a la masticación, esto podría ser ocasionado por procesos tafonómicos, hasta un mal manejo y preservación de los molares.
- Paleodieta en Mammuthus columbi
Aunque Mammuthus columbi ha sido estudiado por mucho tiempo (Ferrusquía-Villafranca, 1978; Gillette y Madsen, 1993; Morett et al., 1998; Saunders, 1999; Siebe et al., 1999; Haynes, 1991; Arroyo-Cabrales et al., 2003; 2007; Maschenko, 2003; Guilbaud et al., 2015; Rochín-Bañaga et al., 2017; Cammidge et al., 2020; Jiménez-Moreno et al., 2021; 2022). El análisis de isotopos estables de carbono y oxígeno en esmalte, de carbono y nitrógeno en colágeno y, el análisis de microdesgaste y mesodesgaste dental han mejorado sustancialmente la manera en que se analiza la alimentación de estos animales, facilitando no solo las reconstrucciones paleodietéticas, sino también la paleoecología y el paleoambiente de los ecosistemas donde habitaron (Fisher et al., 2003; McDaniel y Jefferson, 2006; Pérez-Crespo et al., 2012; Rivals et al., 2012; Gutiérrez-Bedolla; 2014; Saarinen et al., 2015; Metcalfe et al., 2016; Carbot-Chanona et al., 2017; Bravo-Cuevas et al., 2020; Cammidge et al., 2020; Wooller et al., 2021). Mediante el uso de estas herramientas, se ha estimado que este proboscídeo no solamente habitaba en pastizales o ambientes abiertos, sino que han revelado la flexibilidad de sus preferencias alimentarias, con una amplia variación en las plantas que consumían, así mismo, se han podido inferir sus movimientos geográficos e incluso, la estación climática en la que murieron los individuos.
Los estudios paleodietéticos multiproxy, es decir, la combinación de diferentes técnicas para dilucidar la dieta de los organismos, ha demostrado ser la mejor opción para un análisis robusto de la paleodieta en estos organismos, una inferencia dietética mediante una sola técnica, podría muchas veces pasar por alto detalles importantes en el modo en el que se alimentan los organismos, la combinación de diferentes análisis podrían incluso, permitir la estimación de variaciones individuales, estacionales o de hábitat, lo cual podría ser bastante útil en individuos estrictamente generalistas o con diferencias importantes en tiempo geológico (Tütken et al., 2013; Loffredo y DeSantis, 2014; Rivals et al., 2015; Jones et al., 2017; Szabó et al., 2021).
Investigaciones recientes con un enfoque multiproxie han demostrado que la dieta de estos mamuts pudo haber sido aún más compleja de lo que se creía; muchas veces los proxies conservan información de diferentes lapsos de tiempo, esto ayuda a dilucidar la dieta incluso en diferentes momentos de la vida del animal. Por ejemplo, Gutiérrez-Bedolla et al. (2016) realizaron un análisis de isótopos estables de carbono y oxígeno y, de microdesgaste dental, y hallaron diferencias entre los resultados de estas dos técnicas: los resultados del análisis isotópico exhibieron una dieta mixta C3/C4 y el microdesgaste dental una alimentación mayoritariamente de pastos (plantas C4). Por otro lado, Cammidge et al. (2020) encontraron que, en los individuos estudiados, el análisis de mesodesgate dental sugería que una alimentación con preferencia a plantas C3, sin embargo, el análisis de microdesgaste y de cálculos dentales (comúnmente llamado sarro), mostraban una preferencia alimentaria mixta (C3/C4), además, los autores mencionan que los cambios dietéticos que observaron se relacionaba directamente con los cambios de vegetación que ocurrieron en la transición Pleistoceno-Holoceno.
Widga et al. (2020) usando análisis de isotopos estables de carbono y nitrógeno en colágeno y, de carbono y oxígeno en esmalte dental de mamuts, encontraron que estos organismos tuvieron importantes cambios en su dieta a medida que se acercaban periodos con condiciones climáticas glaciares, incorporando mayor cantidad de plantas C3 en su dieta.
Noble et al. (2020), encontraron que existió una influencia de la dinámica glacial en la dieta de M. columbi, mostrando una marcada estacionalidad en su dieta, con una mayor incorporación de plantas C4 en periodos cálidos (Interglacial Sangamonium, hace 125 a 75 mil años AP), comparado con periodos más fríos (Glacial Wisconsin, hace 75 a 11 mil años AP) durante el Pleistoceno tardío.
- Conclusiones
Aunque anteriormente se habían inferido los hábitos alimentarios y el hábitat mediante técnicas de observación, descripción y análisis osteológico, en los últimos años la integración de diferentes metodologías para el análisis del comportamiento dietético, como el análisis del desgaste dental y de las proporciones isotópicas del carbono, nitrógeno y oxígeno en diferentes tejidos mineralizados, ha tenido un importante auge, debido a que los estudios dietéticos no solo se limitan a la alimentación potencial de las especies, sino que también pueden identificarse cambios en su dieta y comportamiento, resultando así estudios más completos, exactos y reales. Sin embargo, muchas veces estos estudios se ven frenados por la falta de ejemplares con las características necesarias para la correcta toma de muestras o mediciones, así como de un buen estado de conservación; adicionalmente, muchas veces los análisis y la adquisición de los equipos no siempre son accesibles.
Asimismo, es importante conocer la biología y comportamiento de la especie que se estudia, así como los límites y alcances de cada proxie estudiado. La presente investigación es un gran ejemplo de la gran complejidad que implica estudiar la paleodieta de una sola especie, ya que la correcta toma de muestras y el correcto análisis de las mismas, es clave para una buena interpretación de los resultados obtenidos, así como la interpretación del ambiente en el que vivieron los animales.
Contribuciones de los Autores
Todos los autores contribuyeron a la conceptualización, metodología, investigación, redacción del manuscrito, elaboración de figuras y revisión del manuscrito.
Financiamiento
Financiamiento PAPIIT #IN101321. Financiamiento por parte de CONACyT, beca para estudios de Doctorado en Ciencias Biológicas de Susana Rodríguez-Franco, CVU: 920655.
Agradecimientos
Agradecemos a los consejos de Arqueología y Paleontología del INAH (401.1S.3-2022/317) y al proyecto “Prehistoria y Paleoambientes en la Cuenca Noroccidental de México” por los permisos para estudiar los especímenes. Gracias también a PAPIIT por financiar este proyecto (#IN101321), al Programa de Posgrado “Posgrado en Ciencias Biológicas”, UNAM, y a CONACyT por apoyar Los estudios de Doctorado en Ciencias Biológicas de Susana Rodríguez-Franco (CVU: 920655). Agradecimientos a la Dra. Marisol Montellano Ballesteros, el Dr. Joaquín Arroyo Cabrales y, a la Dra. Christina I. Barrón-Ortiz, por su apoyo durante los estudios de Doctorado de Susana Rodríguez-Franco. Gracias a Susana Guzmán Gómez (LMF2-LaNaBio), por permitirnos el uso del microscopio multifocal del Instituto de Biología, UNAM, para las microfotografías del microdesgaste dental.
Conflicto de interés
Los autores declaran no tener algun conflicto de interés.
Referencias
Ackermans, N.L., 2020, The history of mesowear: a review: PeerJ, 8, e8519.https://doi.org/10.7717/peerj.8519
Ambrose, S.H., 1991, Effects of diet, climate and physiology on nitrogen isotope abundances in terrestrial foodwebs: Journal of Archaeological Science, 18(3), 293-317. https://doi.org/10.1016/0305-4403(91)90067-Y
Ambrose, S.H., DeNiro, M. J., 1986, The isotopic ecology of East African mammals: Oecologia, 69(3), 395-406. https://doi.org/10.1007/BF00377062
Agenbroad, L.D., 2001, Channel Islands (USA) pygmy mammoths (Mammuthus exilis) compared and contrasted with M. columbi, their continental ancestral stock, in The world of elephants. International Congress, Rome, 473-475.
Agenbroad, L. D., 2003, New absolute dates and comparisons for California’s Mammuthus exilis: Deinsea, 9(1), 1-16.
Arroyo-Cabrales, J., O.J. Polaco, E. Johnson, A.F. Guzmán, 2003, The distribution of the genus Mammuthus in Mexico: Deinsea, 9(1), 27–40.
Arroyo-Cabrales, J., Polaco, O. J., Laurito, C., Johnson, E., Alberdi, M. T., Zamora, A. L. V., 2007, The proboscideans (Mammalia) from Mesoamerica: Quaternary International, 169, 17-23. https://doi.org/10.1016/j.quaint.2006.12.017
Asevedo, L., Winck, G. R., Mothé, D., Avilla, L. S., 2012, Ancient diet of the Pleistocene gomphothere Notiomastodon platensis (Mammalia, Proboscidea, Gomphotheriidae) from lowland mid-latitudes of South America: Stereomicrowear and tooth calculus analyses combined: Quaternary International, 255, 42-52. https://doi.org/10.1016/j.quaint.2011.08.037
Baker, G., Jones, L. H. P., Wardrop, I. D., 1959, Cause of wear in sheeps’ teeth: Nature, 184(4698), 1583-1584. https://doi.org/10.1038/1841583b0
Bakker, E. S., Gill, J. L., Johnson, C. N., Vera, F. W., Sandom, C. J., Asner, G. P., Svenning, J. C., 2016, Combining paleo-data and modern exclosure experiments to assess the impact of megafauna extinctions on woody vegetation: Proceedings of the National Academy of Sciences, 113(4), 847-855. https://doi.org/10.1073/pnas.1502545112
Butler, P.M., 1952, The milk‐molars of Perissodactyla, with remarks on molar occlusion: Proceedings of the Zoological Society of London, 121(4), 777-817. https://doi.org/10.1111/j.1096-3642.1952.tb00784.x
Butler, K., Louys, J., ravouillon, K., 2014, Extending dental mesowear analyses to Australian marsupials, with applications to six Plio-Pleistocene kangaroos from southeast Queensland: Palaeogeography, Palaeoclimatology, Palaeoecology, 408, 11-25. https://doi.org/10.1016/j.palaeo.2014.04.024
Bocherens, H., Drucker, D., 2003, Trophic level isotopic enrichment of carbon and nitrogen in bone collagen: case studies from recent and ancient terrestrial ecosystems: International Journal of Osteoarchaeology, 13(1-2), 46-53. https://doi.org/10.1002/oa.662
Bonafini, M., Pellegrini, M., Ditchfield, P., Pollard, A. M., 2013, Investigation of the ‘canopy effect’ in the isotope ecology of temperate woodlands: Journal of Archaeological Science, 40(11), 3926-3935. https://doi.org/10.1016/j.jas.2013.03.028
Bowen, G.J., Wassenaar, L.I., Hobson, K.A., 2005, Global application of stable hydrogen and oxygen isotopes to wildlife forensics: Oecologia, 143(3), 337-348. https://doi.org/10.1007/s00442-004-1813-y
Bravo-Cuevas, V.M., Ortiz-Caballero, E., Jiménez-Hidalgo, E., Barrón-Ortiz, C.I.,Theodor, J. M., 2020, Taxonomía y hábito alimentario de ejemplares de Mammuthus columbi (Proboscidea: Elephantidae) del centro y sur de México: Boletín de la Sociedad Geológica Mexicana, 72(1), 1-29. https://doi.org/10.18268/bsgm2020v72n1a141019
Britton, K., Grimes, V., Dau, J., Richards, M. P., 2009, Reconstructing faunal migrations using intra-tooth sampling and strontium and oxygen isotope analyses: a case study of modern caribou (Rangifer tarandus granti): Journal of Archaeological Science, 36(5), 1163-1172. https://doi.org/10.1016/j.jas.2009.01.003
Cammidge, T. S., Kooyman, B.,Theodor, J. M., 2020, Diet reconstructions for end-Pleistocene Mammut americanum and Mammuthus based on comparative analysis of mesowear, microwear, and dental calculus in modern Loxodonta africana: Palaeogeography, Palaeoclimatology, Palaeoecology, 538, 109403. https://doi.org/10.1016/j.palaeo.2019.109403
Carbot-Chanona, G., Lagunas-Rodríguez, Z., Jiménez-Moreno, F. J., Suárez, S., 2017, Paleobiological appraisal of two specimens of Pleistocene Mammuthus columbi (Mammalia, Proboscidea, Elephantidae) in Puebla, central Mexico: Boletín de la Sociedad Geológica Mexicana, 69(3), 591-609. https://doi.org/10.18268/bsgm2017v69n3a5
Cerling, T.E., Harris, J.M., 1999, Carbon isotope fractionation between diet and bioapatite in ungulate mammals and implications for ecological and paleoecological studies: Oecologia, 120, 347-363. https://doi.org/10.1007/s004420050868
Cerling, T.E., Harris, J.M., MacFadden, B.J., Leakey, M.G., Quade, J., Eisenmann, V., Ehleringer, J.R., 1997, Global vegetation change through the Miocene/Pliocene boundary: Nature, 389(6647), 153-158. https://doi.org/10.1038/38229
Cerling, T.E., Harris, J.M., Leakey, M.G., 1999, Browsing and grazing in elephants: the isotope record of modern and fossil proboscideans: Oecologia, 120(3), 364-374. https://doi.org/10.1007/s004420050869
Cerling, T.E., Harris, J.M., Leakey, M.G., 2005, Environmentally Driven Dietary Adaptations in African Mammals, in Baldwin, I.T.,Caldwell, M.M.,Heldmaier, G.,Jackson, R.B, Lange, O.L.,Mooney, H.A.,Schulze, E.D. Sommer, U.,Ehleringer, J.R., Dearing, M.D.,Cerling, T.E.,(Eds.), A History of Atmospheric CO2 and Its Effects on Plants, Animals, and Ecosystems. Ecological Studies, vol 177: Springer, New York, 258-292. https://doi.org/10.1007/0-387-27048-5_12
Cherney, M. D., Fisher, D. C., Hren, M. T., Shirley, E. A., 2021, Stable isotope records of nursing and weaning: A case study in elephants with implications for paleobiological investigations: Palaeogeography, Palaeoclimatology, Palaeoecology, 567, 110223. https://doi.org/10.1016/j.palaeo.2021.110223
Codron, J., Codron, D., Lee-Thorp, J.A., Sponheimer, M., Kirkman, K., Duffy, K. J., Sealy, J., 2011, Landscape-scale feeding patterns of African elephant inferred from carbon isotope analysis of feces: Oecologia, 165(1), 89-99. https://doi.org/10.1007/s00442-010-1835-6
Danowitz, M., Hou, S., Mihlbachler, M., Hastings, V., Solounias, N., 2016, A combined-mesowear analysis of late Miocene giraffids from North Chinese and Greek localities of the Pikermian Biome: Palaeogeography, Palaeoclimatology, Palaeoecology, 449, 194-204. https://doi.org/10.1016/j.palaeo.2016.02.026
Dansgaard, W., 1964, Stable isotopes in precipitation: Tellus, 16, 436–468. https://doi.org/10.3402/tellusa.v16i4.8993
Delwiche, C.C., Zinke, P.J., Johnson, C.M., Virginia, R.A.,1979, Nitrogen isotope distribution as a presumptive indicator of nitrogen fixation: Botanical Gazette, 140, S65-S69. https://doi.org/10.1086/337037
DeNiro, M. J., Epstein, S., 1981, Influence of diet on the distribution of nitrogen isotopes in animals: Geochimica et cosmochimica acta, 45(3), 341-351. https://doi.org/10.1016/0016-7037(81)90244-1
DeSantis, L.R., 2016, Dental microwear textures: reconstructing diets of fossil mammals: Surface Topography: Metrology and Properties, 4(2), 023002. https://doi.org/10.1088/2051-672X/4/2/023002
Dutton, A., Lohmann, K.C., Leckie, R.M., 2005, Insights from the Paleogene tropical Pacific: Foraminiferal stable isotope and elemental results from Site 1209, Shatsky Rise: Paleoceanography, 20(3), PA3004. https://doi.org/10.1029/2004PA001098
Edwards, E.J., Osborne, C.P., Strömberg, C. A., Smith, S. A., C4 Grasses Consortium, Bond, W.J., Pascal-Antoine, C., Cousins, A.B., Duvall, M., Fox, D.L., Freckleton, R.P., Ghannoum, O., Hartwell, J., Huang, Y., Janis, C.M., Keeley, J.E., Kellogg, E.A., Knapp, A.K., Leakey, A.D.B., Nelson, D.M., Saarela, J.M., Sage, R.F., Sala, O.E., Salamin, N., Still, C.J., Tipple, B., 2010, The origins of C4 grasslands: integrating evolutionary and ecosystem science: Science. 328(5978), 587-591. https://doi.org/10.1126/science.1177216
Ehleringer, J.R., Monson, R.K., 1993, Evolutionary and ecological aspects of photosynthetic pathway variation: Annual Review of Ecology and Systematics, 24(1), 411-439. https://doi.org/10.1146/annurev.es.24.110193.002211
Epstein, S., Buchsbaum, R., Lowenstam, H., Urey, H. C., 1951, Carbonate-waterisotopic temperatura scale: Geological Society of America Bulletin, 62(4), 417-426. https://doi.org/10.1130/0016-7606(1951)62[417:CITS]2.0.CO;2
Epstein, S., Mayeda, T., 1953, Variation of 18O content of waters from natural sources: Geochimica et Cosmochimica Acta, 4(5), 213-224. https://doi.org/10.1016/0016-7037(53)90051-9
Farquhar, G.D., Ehleringer, J.R., Hubrick, K. T., 1989, Carbon isotopic discrimination and photosynthesis: Annual Review of Plant Physiology and Plant Molecular Biology, 40, 503–537. https://doi.org/10.1146/annurev.pp.40.060189.002443
Fernández-García, M., López-García, J.M., Royer, A., Lécuyer, C., Rivals, F., Rufà, A., Blasco, R., Rosell, J., 2022, New insights in Neanderthal palaeoecology using stable oxygen isotopes preserved in small mammals as palaeoclimatic tracers in Teixoneres Cave (Moià, northeastern Iberia): Archaeological and Anthropological Sciences, 14(6), 1-25. https://doi.org/10.1007/s12520-022-01564-9
Ferretti, M.P., 2003, Structure and evolution of mammoth molar enamel: Acta Palaeontologica Polonica, 48(3), 383–396.
Ferrusquía-Villafranca, I., 1978, Distribution of Cenozoic vertebrate faunas in Middle America and problems of migration between North and South America, en Ferrusquía-Villafranca, I., (Ed.), Conexiones terrestres entre Norte y Sudamérica: México, Universidad Nacional Autónoma de México, Boletín del Instituto de Geología, 193-321.
Fisher, D.C., Fox, D.L., Agenbroad, L.D., 2003, Tusk growth rate and season of death of Mammuthus columbi from Hot Springs, South Dakota, USA: Deinsea, 9(1), 117–134.
Fortelius, M., 1985, Ungulate cheek teeth: developmental, functional, and evolutionary interrelations: Acta Zoologica Fennica, 180, 1-76.
Fortelius, M., Solounias, N., 2000, Functional characterization of ungulate molars using the abrasion-attrition wear gradient: a new method for reconstructing paleodiets: American Museum Novitates, 2000(3301), 1-36. https://doi.org/10.1206/0003-0082(2000)301%3C0001:FCOUMU%3E2.0.CO;2
Fox, D.L., Fisher, D.C., 2004, Dietary reconstruction of Miocene Gomphotherium (Mammalia, Proboscidea) from the Great Plains region, USA, based on the carbon isotope composition of tusk and molar enamel: Palaeogeography, Palaeoclimatology, Palaeoecology, 206(3-4), 311-335. https://doi.org/10.1016/j.palaeo.2004.01.010
Fox-Dobbs, K., Leonard, J.A., Koch, P.L., 2008, Pleistocene megafauna from eastern Beringia: paleoecologic and paleoenvironmental interpretations of stable carbon and nitrogen isotope and radiocarbon records: Palaeogeography, Palaeoclimatology, Palaeoecology, 21, 30-46. https://doi.org/10.1016/j.palaeo.2007.12.011
Fraser, D., Mallon, J. C., Furr, R., Theodor, J. M., 2009, Improving the repeatability of low magnification microwear methods using high dynamic range imaging: Palaios, 24(12), 818-825. https://doi.org/10.2110/palo.2009.p09-064r
Fraser, D., Theodor, J. M, 2011, Comparing ungulate dietary proxies using discriminant function analysis: Journal of Morphology, 272(12), 1513-1526. https://doi.org/10.1002/jmor.11001
Gillette, D. D., Madsen, D. B., 1993, The Columbian mammoth, Mammuthus columbi, from the Wasatch Mountains of central Utah: Journal of Paleontology, 67(4), 669-680. https://doi.org/10.1017/S0022336000024999
González-Guarda, E., Petermann-Pichincura, A., Tornero, C., Domingo, L., Agustí, J., Pino, M., Abarzúa, A.M., Villavicencio, N.A., Labarca, R., Tolorza, V., Sevilla, P., Rivals, F., 2018, Multiproxy evidence for leaf-browsing and closed habitats in extinct proboscideans (Mammalia, Proboscidea) from Central Chile: Proceedings of the National Academy of Sciences, 115(37), 9258-9263. https://doi.org/10.1073/pnas.1804642115
Gordon, K.D., 1982, A study of microwear on chimpanzee molars: implications for dental microwear análisis: American Journal of Physical Anthropology, 59(2), 195-215. https://doi.org/10.1002/ajpa.1330590208
Gowik, U., Westhoff, P., 2011, The path from C3 to C4 photosynthesis: Plant Physiology, 155(1), 56-63. https://doi.org/10.1104/pp.110.165308
Green, J.L., Croft, D.A., 2018, Using dental mesowear and microwear for dietary inference: a review of current techniques and applications: Methods in Paleoecology, 53-73. https://doi.org/10.1007/978-3-319-94265-0_5
Greyling, M.D., 2004, Sex and age related distinctions in the feeding ecology of the African elephant Loxodonta africana: Witwatersrand, Sudafrica, Universidad de Witwatersrand, tesis doctoral, 189 p.
Grigoriev, S., Tikhonov, A., Fisher, D., Obada, T., Savvinov, G., Novgorodov, G., Cheprasov, M., Fedorov, S., Ivanov, E., 2014, Discovery of a woolly mammoth (Mammuthus primigenius) carcass from Malyi Lyakhovski Island (New Siberian Islands), in VI International Conference on Mammoths and Their Relatives, Scientific Annals of the School of Geology, Vol. 102, 64-65.
Grigoriev, S.E., Fisher, D.C., Obadă, T., Shirley, E.A., Rountrey, A.N., Savvinov, G.N., Garmaeva, D.K., Novgorodov, G.P., Cheprasov, M.Y., Vasilev, S.E., Goncharov, A.E., Masharskiy, A., Egorova, V.E., Petrova, P.P., Egorova, E.E., Akheremenko, Y.A., van der Plicht, J., Galanin, A.A., Federov, S.E., Ivanov, E.V., Tikhonov, A. N., 2017, A woolly mammoth (Mammuthus primigenius) carcass from maly Lyakhovsky island (new siberian islands, Russian Federation): Quaternary International, 445, 89-103. https://doi.org/10.1016/j.quaint.2017.01.007
Grine, F.E., 1986, Dental evidence for dietary differences in Australopithecus and Paranthropus: a quantitative analysis of permanent molar microwear: Journal of human evolution, 15(8), 783-822. https://doi.org/10.1016/S0047-2484(86)80010-0
Grine, F. E., Ungar, P. S., Teaford, M. F., 2002, Error rates in dental microwear quantification using scanning electron microscopy: Scanning: The Journal of Scanning Microscopies, 24(3), 144-153. https://doi.org/10.1002/sca.4950240307
Guilbaud, M.N., Arana-Salinas, L., Siebe, C., Barba-Pingarrón, L.A., Ortiz, A., 2015, Volcanic stratigraphy of a high-altitude Mammuthus columbi (Tlacotenco, sierra Chichinautzin), central México: Bulletin of Volcanology, 77(3), 1-16. https://doi.org/10.1007/s00445-015-0903-
Gutiérrez-Bedolla, M., García-Zepeda, M.L., López-García, R., Arroyo-Cabrales, J., Marín-Leyva, A.H., Meléndez-Herrera, E., Fuentes-Farías, A. L., 2016, Diet and habitat of Mammuthus columbi (Falconer, 1857) from two Late Pleistocene localities in central western Mexico: Quaternary International. 406, 137-146. https://doi.org/10.1016/j.quaint.2015.08.022
Haynes, G., 1991, Mammoths, Mastodons, and Elephants: England, Cambridge University Press, 413 p.
Haynes, G., 1993, Mammoths, mastodonts, and elephants: biology, behavior and the fossil record: Nwe York, NY, Cambridge University Press, 56-59.
Haynes, G., 2012, Elephants (and extinct relatives) as earth-movers and ecosystem engineers: Geomorphology, 157, 99-107. https://doi.org/10.1016/j.geomorph.2011.04.045
Haynes, G., Klimowicz, J., 2015, A preliminary review of bone and teeth abnormalities seen in recent Loxodonta and extinct Mammuthus and Mammut, and suggested implications: Quaternary International, 379, 135-146. https://doi.org/10.1016/j.quaint.2015.04.001
Heaton, T. H., 1987, The 15N/14N ratios of plants in South Africa and Namibia: relationship to climate and coastal/saline environments: Oecologia, 74, 236-246.
Hedges, R. E., 2003, On bone collagen-apatite-carbonate isotopic relationships: International Journal of Osteoarchaeology, 13(1‐2), 66-79. https://doi.org/10.1002/oa.660
Higgins, P., 2018, Isotope ecology from biominerals, in Croft D., Su, D., Simpson, S. (Eds.) Methods in Paleoecology Vertebrate Paleobiology and Paleoanthropology. Cham, Springer. 99-120. https://doi.org/10.1007/978-3-319-94265-0_
Hillson, S., 2005, Teeth: UK, Cambridge University Press, 373 p.
Hobson, K.A., Bairlein, F., 2003, Isotopic fractionation and turnover in captive Garden Warblers (Sylvia borin): implications for delineating dietary and migratory associations in wild passerines: Canadian Journal of Zoology, 81(9), 1630-1635. https://doi.org/10.1139/z03-140
Hoffman, J. M., Fraser, D., Clementz, M. T., 2015, Controlled feeding trials with ungulates: a new application of in vivo dental molding to assess the abrasive factors of microwear: The Journal of Experimental Biology, 218(10), 1538-1547. https://doi.org/10.1242/jeb.118406
Iacumin, P., Di Matteo, A., Nikolaev, V., Kuznetsova, T. V., 2010, Climate information from C, N and O stable isotope analyses of mammoth bones from northern Siberia: Quaternary International, 212(2), 206-212. https://doi.org/10.1016/j.quaint.2009.10.009
ImageJ, 2022, ImageJ Image Processing and Analysis in Java, versión 1.53: https://imagej.nih.gov/ij/docs/index.html
Janis, C.M., Fortelius, M., 1988, On the means whereby mammals achieve increased functional durability of their dentitions, with special reference to limiting factors: Biological Reviews, 63(2), 197-230. https://doi.org/10.1111/j.1469-185X.1988.tb00630.x
Jaouen, K., Villalba-Mouco, V., Smith, G.M., Trost, M., Leichliter, J., Lüdecke, T., Méjean, P., Mandrou, S., Chmeleff, J., Guiserix, D., Bourgon, N., Blasco F., Cardoso, J.M., Duquenoy, C., Moubtahij, Z., Garcia, D.C.S., Richards, M., Tüken, T., Hublin, J., Utrilla, P., Montes, L., 2022, A Neandertal dietary conundrum: Insights provided by tooth enamel Zn isotopes from Gabasa, Spain: Proceedings of the National Academy of Sciences, 119(43), e2109315119. https://doi.org/10.1073/pnas.2109315119
Jenkins, S. G., Partridge, S. T., Stephenson, T. R., Farley, S. D., Robbins, C. T., 2001, Nitrogen and carbon isotope fractionation between mothers, neonates, and nursing offspring: Oecologia, 129(3), 336-341. https://doi.org/10.1007/s004420100755
Jiménez-Manchón, S., Blaise, É., Albesso, M., Gardeisen, A., Rivals, F., 2022, Quantitative dental mesowear analysis in domestic Caprids: a new method to reconstruct management strategies: Journal of Archaeological Method and Theory, 29(2), 540-560. https://doi.org/10.1007/s10816-021-09530-w
Jiménez-Moreno, F. J., Morales-Tehuitzitl, E.D., Carbot-Chanona, G., Velázquez-Castro, J., 2022, A mathematical model to calculate the population of Mammuthus columbi (Mammalia, Proboscidea, Elephantidae) during the Late Pleistocene in the Valsequillo Basin, Puebla, Mexico: Historical Biology, 34(4), 750-758. https://doi.org/10.1080/08912963.2021.1946530
Jones, D. B., DeSantis, L. R., 2017, Dietary ecology of ungulates from the La Brea tar pits in southern California: a multi-proxy approach: Palaeogeography, Palaeoclimatology, Palaeoecology, 466, 110-127. https://doi.org/10.1016/j.palaeo.2016.11.019
Kaiser, T. M., Müller, D. W., Fortelius, M., Schulz, E., Codron, D., Clauss, M., 2013, Hypsodonty and tooth facet development in relation to diet and habitat in herbivorous ungulates: implications for understanding tooth wear: Mammal Review, 43(1), 34-46. https://doi.org/10.1111/j.1365-2907.2011.00203.x
Kaiser, T. M., Solounias, N., Fortelius, M., Bernor, R. L., Schrenk, F., 2000, Tooth mesowear analysis on Hippotheriun primigenium from the Vallesian Dinotheriensande (Germany) — a blind test study: Carolinea, 58, 103–114.
Kaiser, T., Solounias, N., 2003, Extending the tooth mesowear method to extinct and extant equids: Geodiversitas, 25, 321–345.
Keeling, R.F., Graven, H.D., Welp, L.R., Resplandy, L., Bi, J., Piper, S.C., Sun, Y., Bollenbacher, A., Meijer, H.A., 2017, Atmospheric evidence for a global secular increase in carbon isotopic discrimination of land photosynthesis: Proceedings of the National Academy of Sciences, 114(39), 10361-10366. https://doi.org/10.1073/pnas.1619240114
Kelly, J.F., 2000, Stable isotopes of carbon and nitrogen in the study of avian and mammalian trophic ecology: Canadian Journal of Zoology, 78(1), 1-27. https://doi.org/10.1139/z99-165
Koch, P.L., 1994, Tracing the diets of fossil animals using stable isotopes, in Lajtha, K., Michener, R. (eds.), Stable isotopes in ecology and environmental science: Boston, Blackwell Scientific Publication, 63-92.
Koch, P.L., 1998, Isotopic reconstruction of past continental environments: Annual Review Earth Planetary Science, 26, 573-613. https://doi.org/10.1146/annurev.earth.26.1.573
Koch, P. L., 2007, Isotopic study of the biology of modern and fossil vertebrates, in Michener, R., Lajtha, K., (eds.), Stable isotopes in ecology and environmental science: Boston, Blackwell Publishing, 99-154
Koch, P.L., Fisher, D.C., Dettman, D., 1989, Oxygen isotope variation in the tusks of extinct proboscideans: a measure of season of death and seasonality: Geology, 17(6), 515-519. https://doi.org/10.1130/0091-7613(1989)017%3C0515:OIVITT%3E2.3.CO;2
Kohn, M.J., 1996, Predicting animal δ18O: accounting for diet and physiological adaptation: Geochimica et Cosmochimica Acta, 60(23), 4811-4829. https://doi.org/10.1016/S0016-7037(96)00240-2
Kohn, M. J., 2010, Carbon isotope compositions of terrestrial C3 plants as indicators of (paleo) ecology and (paleo) climate: Proceedings of the National Academy of Sciences, 107(46), 19691-19695. https://doi.org/10.1073/pnas.1004933107
Koirala, R.K., Raubenheimer, D., Aryal, A., Pathak, M. L., Ji, W., 2016, Feeding preferences of the Asian elephant (Elephas maximus) in Nepal: BMC Ecology, 16(1), 1-9. https://doi.org/10.1186/s12898-016-0105-9
Krajcarz, M. T., Krajcarz, M., Bocherens, H., 2018, Collagen-to-collagen prey-predator isotopic enrichment (Δ13C, Δ15N) in terrestrial mammals-a case study of a subfossil red fox den: Palaeogeography, Palaeoclimatology, Palaeoecology, 490, 563-570. https://doi.org/10.1016/j.palaeo.2017.11.044
Krylovich, O. A., Boeskorov, G. G., Shchelchkova, M. V., Savinetsky, A. B., 2020, The trophic position of Pleistocene and modern brown bears (Ursus arctos) of Yakutia based on stable isotope analyses: Biology Bulletin, 47(8), 1013-1021. https://doi.org/10.1134/S1062359020080087
Krzemińska, A., 2008, Preliminary characteristics of pathologies found in the skeletons of mammoths at the kraków spadzista street (b) site: Veterinarija ir Zootechnika, 43(65), 52-57.
Laws, R. M., 1966, Age criteria for the African elephant, Loxodonta A. Africana: East African Wildlife Journal, 4, 1-37. https://doi.org/10.1111/j.1365-2028.1966.tb00878.x
LeGeros, R.Z., 1991,Calcium phosphates in oral biology and medicine, Vol 15: Basel, Karger, 201p. https://doi.org/10.1159/isbn.978-3-318-04021-0
Lister, A. M., 2014, Behavioural leads in evolution: evidence from the fossil record: Biological Journal of the Linnean Society, 112(2), 315-331. https://doi.org/10.1111/bij.12173
Lister, A.M., Sher, A.V., 2015, Evolution and dispersal of mammoths across the Northern Hemisphere: Science, 350(6262), 805-809. https://doi.org/10.1126/science.aac5660
Liu, Z., Prendergast, A., Drysdale, R., May, J. H., 2022, Comparison of bulk and sequential sampling methodologies on mammoth tooth enamel and their implications in paleoenvironmental reconstructions: E&G Quaternary Science Journal, 71(2), 227-241. https://doi.org/10.5194/egqsj-71-227-2022
Loffredo, L.F., DeSantis, L.R., 2014, Cautionary lessons from assessing dental mesowear observer variability and integrating paleoecological proxies of an extreme generalist Cormohipparion emsliei: Palaeogeography, Palaeoclimatology, Palaeoecology, 395, 42-52. https://doi.org/10.1016/j.palaeo.2013.12.020
Longinelli, A., 1984, Oxygen isotopes in mammal bone phosphate: ¿a new tool for paleohydrological and paleoclimatological research?: Geochimica et Cosmochimica Acta, 48(2), 385-390. https://doi.org/10.1016/0016-7037(84)90259-X
Lowenstam, H.A. Weiner, S., 1989, On biomineralization: UK, Oxford University Press, 324p. https://doi.org/10.1093/oso/9780195049770.001.0001
Lundgren, E. J., Schowanek, S.D., Rowan, J., Middleton, O., Pedersen, R.Ø., Wallach, A.D., Ramp, D., Davis, M., Sandom, C.J., Svenning, J.C., 2021, Functional traits of the world’s late Quaternary large-bodied avian and mammalian herbivores: Scientific Data, 8(1), 1-21. https://doi.org/10.6084/m9.figshare.13353416
MacFadden, B.J., Cerling, T. E., 1996, Mammalian herbivore communities, ancient feeding ecology, and carbon isotopes: a 10 million year sequence from the Neogene of Florida: Journal of Vertebrate Paleontology. 16(1), 103-115. https://doi.org/10.1080/02724634.1996.10011288
Mainland, I. L., 2003, Dental microwear in grazing and browsing Gotland sheep (Ovis aries) and its implications for dietary reconstruction: Journal of Archaeological Science. 30(11), 1513-1527. https://doi.org/10.1016/S0305-4403(03)00055-4
Martínez-Polanco, M.F., Ramírez-Pedraza, I., Guillén-Espínola, M.D., Rivals, F., 2021, Protocol for molding and casting teeth for microwear análisis: Zenodo, (Video). https://doi.org/10.5281/zenodo.4926101
Maschenko, E., 2002, Individual development, biology and evolution of the woolly mammoth: Cranium 19 (1), 4–120.
McComb, K., Moss, C., Durant, S. M., Baker, L., Sayialel, S., 2001, Matriarchs as repositories of social knowledge in African elephants: Science, 292(5516), 491-494. https://doi.org/10.1126/science.1057895
McDaniel Jr, G.E., Jefferson, G.T., 2006, Dental variation in the molars of Mammuthus columbi var. M. imperator (Proboscidea, Elephantidae) from a Mathis gravel quarry, southern Texas: Quaternary International, 142, 166-177. https://doi.org/10.1016/j.quaint.2005.03.014
Medrano, H., Flexas, J., 2000, Fotorrespiración y mecanismos de concentración del dióxido de carbono, en Ázcón-Bieto, J., Talón, M.,(eds.), Fundamentos de fisiología vegetal: Madrid, McGraw-Hill Interamericana, 187-201.
Metcalfe, J.Z., 2017, Proboscidean isotopic compositions provide insight into ancient humans and their environments: Quaternary International, 443, 147-159. https://doi.org/10.1016/j.quaint.2016.12.003
Metcalfe, J.Z., Longstaffe, F.J., Zazula, G. D., 2010, Nursing, weaning, and tooth development in woolly mammoths from Old Crow, Yukon, Canada: implications for Pleistocene extinctions: Palaeogeography, Palaeoclimatology, Palaeoecology, 298(3-4), 257-270. https://doi.org/10.1016/j.palaeo.2010.09.032
Metcalfe, J. Z., Longstaffe, F. J., Hodgins, G., 2013, Proboscideans and paleoenvironments of the Pleistocene Great Lakes: landscape, vegetation, and stable isotopes: Quaternary Science Reviews, 76, 102-113. https://doi.org/10.1016/j.quascirev.2013.07.004
Mihlbachler, M. C., Solounias, N., 2006, Coevolution of tooth crown height and diet in oreodonts (Merycoidodontidae, Artiodactyla) examined with phylogenetically independent contrasts: Journal of Mammalian Evolution, 13(1), 11-36. https://doi.org/10.1007/s10914-005-9001-3
Mihlbachler, M.C., Rivals, F., Solounias, N., Semprebon, G.M., 2011, Dietary change and evolution of horses in North America: Science, 331(6021), 1178-1181. https://doi.org/10.1126/science.1196166
Miller, J. H., Fisher, D. C., Crowley, B. E., Secord, R., Konomi, B. A., 2022, Male mastodon landscape use changed with maturation (late Pleistocene, North America): Proceedings of the National Academy of Sciences, 119(25), e2118329119. https://doi.org/10.1073/pnas.2118329119
Mills, J. R. E., 1955, Ideal dental occlusion in the primates: Dent Practit, 6(2), 47-61.
Morales-Puente, P., Cienfuegos, E., Manzanilla, L.R., Otero, F.J., 2012, Estudio de la paleodieta empleando isótopos estables de los elementos carbono, oxígeno y nitrógeno en restos humanos y fauna encontrados en el barrio teotihuacano de Teopancazco, Teotihuacan, en Manzanilla, L., R. (ed.), Estudios arqueométricos del centro de barrio de Teopancazco en Teotihuacan: Mexico, Uiversidad Nacional Autónoma de México. 347-422.
Morett, L.A., Arroyo-Cabrales, J., Polaco, O. J., 1998, Tocuila, a remarkable mammoth site in the Basin of Mexico: Current Research in the Pleistocene, 15, 118-120.
Moss, C., 2012, Elephant memories: thirteen years in the life of an elephant family: USA, University of Chicago Press, 402 p.
Muhs, D.R., Simmons, K.R., Groves, L.T., McGeehin, J.P., Schumann, R.R., Agenbroad, L.D., 2015, Late Quaternary sea-level history and the antiquity of mammoths (Mammuthus exilis and Mammuthus columbi), Channel Islands National Park, California, USA: Quaternary Research, 83(3), 502-521. https://doi.org/10.1016/j.yqres.2015.03.001
Noble, E. J., McManus, J. G., Mead, A. J., Mead, H., Seminack, C., Balco, W., Bennett, T., Crain, N.M., Duckworth, C., Malasek, T., Pearso, J.Z., Rhinehart, P., Ussery, M.E., Sun, Y., Patterson, J.R., Patterson, D.B., 2020, Enamel isotopes reveal late Pleistocene ecosystem dynamics in southeastern North America: Quaternary Science Reviews, 236, 106284. https://doi.org/10.1016/j.quascirev.2020.106284
Owen-Smith, N., 1987, Pleistocene extinctions: the pivotal role of megaherbivores: Paleobiology, 13(3), 351-362. https://doi:10.1017/S0094837300008927
Pappa, S., Schreve, D. C., Rivals, F., 2019, The bear necessities: A new dental microwear database for the interpretation of palaeodiet in fossil Ursidae: Palaeogeography, Palaeoclimatology, Palaeoecology, 514, 168-188. https://doi.org/10.1016/j.palaeo.2018.10.015
Parkman, B., 2002, Mammoth rocks: where Pleistocene giants got a good rub? Mammoth Trumpet, 18(1),4-7,
Parkman, B., 2009, Rubbing Rocks, Vernal Pools, and the First Californians: Pursuing the Rancholabrean hypothesis.
Pérez-Crespo, V.A., Arroyo-Cabrales, J., Benammi, M., Johnson, E., Polaco, O.J., Santos-Moreno, A., Morales-Puente, P., Cienfuegos-Alvarado, E., 2012, Geographic variation of diet and habitat of the Mexican populations of Columbian Mammoth (Mammuthus columbi): Quaternary International. 276, 8-16. https://doi.org/10.1016/j.quaint.2012.03.014
Ramírez-Pedraza, I., Tornero, C., Pappa, S., Talamo, S., Salazar-García, D. C., Blasco, R., Rosell, J., Rivals, F., 2019, Microwear and isotopic analyses on cave bear remains from Toll Cave reveal both short-term and long-term dietary habits: Scientific Reports, 9(1), 5716. https://doi.org/10.1038/s41598-019-42152-7
Raubenheimer, E. J., 2000, Development of the tush and tusk and tusklessness in African elephant (Loxodonta africana): Koedoe, 43(2), 57-64. https://doi.org/10.4102/koedoe.v43i2.199
Raubenheimer, E. J., Van Heerden, W. F. P., Van Niekerk, P. J., De Vos, V., Turner, M. J., 1995, Morphology of the deciduous tusk (tush) of the African elephant (Loxodonta africana): Archives of Oral Biology, 40(6), 571-576. https://doi.org/10.1016/0003-9969(95)00008-D
Rensberger, J.M., 1978, Scanning electron microscopy of wear and occlusal events in some small herbivores, in Teaford, M.F., Smith, M.M., Ferguson, M.W., (eds.), Development, function and evolution of teeth: UK, Cambridge University Press, 415-438.
Rivals, F., Baryshnikov, G. F., Prilepskaya, N. E., Belyaev, R. I., 2022, Diet and ecological niches of the Late Pleistocene hyenas Crocuta spelaea and C. ultima ussurica based on a study of tooth microwear: Palaeogeography, Palaeoclimatology, Palaeoecology, 601, 111125. https://doi.org/10.1016/j.palaeo.2022.111125
Rivals, F., Julien, M.A., Kuitems, M., Van Kolfschoten, T., Serangeli, J., Drucker, D. G., Bocherens, H, Conard, N.J., 2015, Investigation of equid paleodiet from Schöningen 13 II-4 through dental wear and isotopic analyses: Archaeological implications: Journal of Human Evolution, 89, 129-137. https://doi.org/10.1016/j.jhevol.2014.04.002
Rivals, F., Mihlbachler, M.C., Solounias, N., 2007, Effect of ontogenetic-age distribution in fossil and modern samples on the interpretation of ungulate paleodiets using the mesowear method: Journal of Vertebrate Paleontology, 27(3), 763-767. https://doi.org/10.1671/0272-4634(2007)27[763:EOODIF]2.0.CO;2
Rivals, F., Rindel, D., Belardi, J. B., 2013, Dietary ecology of extant guanaco (Lama guanicoe) from Southern Patagonia: seasonal leaf browsing and its archaeological implications: Journal of Archaeological Science, 40(7), 2971-2980. https://doi.org/10.1016/j.jas.2013.03.005
Rivals, F., Semprebon, G., Lister, A., 2012, An examination of dietary diversity patterns in Pleistocene proboscideans (Mammuthus, Palaeoloxodon, and Mammut) from Europe and North America as revealed by dental microwear: Quaternary International, 255, 188-195. https://doi.org/10.1016/j.quaint.2011.05.036
Rivals, F., Semprebon, G.M., Lister, A.M., 2019, Feeding traits and dietary variation in Pleistocene proboscideans: A tooth microwear review: Quaternary Science Reviews. 219, 145-153. https://doi.org/10.1016/j.quascirev.2019.06.027
Rochín-Bañaga, H., Schwennicke, T., Herrera-Gil, L. A., 2017, Registro de Mammuthus columbi en el área de El Carrizal, Baja California Sur, México: Boletín de la Sociedad Geológica Mexicana, 69(2), 495-503 . https://doi.org/10.18268/bsgm2017v69n2a12
Roth, V.L., 1989, Fabricational noise in elephant dentitions: Paleobiology, 15(2), 165-179. https://doi.org/10.1017/S0094837300009349
Roth, V.L.,1992, Quantitative variation in elephant dentitions: implications for the delimitation of fossil species: Paleobiology, 18(2), 184-202. https://doi.org/10.1017/S0094837300013968
Royer, A., Lécuyer, C., Montuire, S., Escarguel, G., Fourel, F., Mann, A., Maureille, B., 2013, Late Pleistocene (MIS 3–4) climate inferred from micromammal communities and δ18O of rodents from Les Pradelles, France: Quaternary Research, 80(1), 113-124. https://doi.org/10.1016/j.yqres.2013.03.007
Royer, A., Lécuyer, C., Montuire, S., Primault, J., Fourel, F., Jeannet, M., 2014, Summer air temperature, reconstructions from the last glacial stage based on rodents from the site Taillis-des-Coteaux (Vienne), Western France: Quaternary Research, 82(2), 420-429. https://doi.org/10.1016/j.yqres.2014.06.006
Rountrey, A.N., Fisher, D.C., Tikhonov, A.N., Kosintsev, P.A., Lazarev, P.A., Boeskorov, G., Buigues, B., 2012, Early tooth development, gestation, and season of birth in mammoths: Quaternary International, 255, 196-205. https://doi.org/10.1016/j.quaint.2011.06.006
Rozanski, K., Araguás-Araguás, L., Gonfiantini, R., 1993, Isotopic patterns in modern global precipitation: Climate Change in Continental Isotopic Records, 78: USA, American Geophysical Union, 1-36. https://doi.org/10.1029/GM078p0001
Saarinen, J., Karme, A., Cerling, T., Uno, K., Säilä, L., Kasiki, S., Ngene, S., Obari, T.,Mbua, E., Manthi, F.K., Fortelius, M., 2015, A new tooth wear–based dietary analysis method for Proboscidea (Mammalia): Journal of Vertebrate Paleontology, 35 (3), e918546. https://doi.org/10.1080/02724634.2014.918546
Saarinen, J., Karme, A., 2017, Tooth wear and diets of extant and fossil xenarthrans (Mammalia, Xenarthra)–applying a new mesowear approach: Palaeogeography, Palaeoclimatology, Palaeoecology, 476, 42-54. https://doi.org/10.1016/j.palaeo.2017.03.027
Saarinen, J., Karme, A., Cerling, T., Uno, K., Säilä, L., Kasiki, S., Ngene, S., Obari, T. ,Mbua, E., Manthi, F.K., Fortelius, M., 2015, A new tooth wear–based dietary analysis method for proboscidea (mammalia): Journal of Vertebrate Paleontology, 35 (3), e918546. https://doi.org/10.1080/02724634.2014.918546
Sanson, G., 2006, The biomechanics of browsing and grazing: American Journal of Botany, 93(10), 1531-1545. https://doi.org/10.3732/ajb.93.10.1531
Saunders, J. J., 1999, Morphometrical analyses of Mammuthus columbi from the Dent site, Weld county, Colorado: Deinsea, 6(1), 55-78.
Schoeninger, M., DeNiro, M.J., 1984, Nitrogen and carbon isotopic composition of bone collagen from marine and terrestrial animals: Geochimica et Cosmochimica Acta, 48, 625-639. https://doi.org/10.1016/0016-7037(84)90091-7
Schoeninger, M.J., Moore, K.M., Murray, M.L., Kingston, J.D., 1989, Detection of bone preservation in archaeological and fossil simples: Applied Geochemistry, 4(3), 281-292. https://doi.org/10.1016/0883-2927(89)90030-9
Schulte, B. A., 2000, Social structure and helping behavior in captive elephants. Zoo Biology, 19(5), 447-459. https://doi.org/10.1002/1098-2361(2000)19:5%3C447::AID-ZOO12%3E3.0.CO;2-%23
Scott, R. S., Ungar, P. S., Bergstrom, T. S., Brown, C. A., Grine, F. E., Teaford, M. F., Walker, A., 2005, Dental microwear texture analysis shows within-species diet variability in fossil hominins: Nature, 436(7051), 693-695. https://doi.org/10.1038/nature03822
Scott, R. S., Ungar, P. S., Bergstrom, T. S., Brown, C. A., Childs, B. E., Teaford, M. F., Walker, A., 2006, Dental microwear texture analysis: technical considerations: Journal of Human Evolution, 51(4), 339-349. https://doi.org/10.1016/j.jhevol.2006.04.006
Schwartz-Narbonne, R., Longstaffe, F. J., Kardynal, K. J., Druckenmiller, P., Hobson, K.A., Jass, C. N., Metcalfe, J.Z., Zazula, G., 2019, Reframing the mammoth steppe: insights from analysis of isotopic niches: Quaternary Science Reviews, 215, 1-21. https://doi.org/10.1016/j.quascirev.2019.04.025
Sealy, J.C., Van Der Merwe, N. J., Thorp, J. A.L., Lanham, J.L., 1987, Nitrogen isotopic ecology in southern Africa: implications for environmental and dietary tracing: Geochimica et Cosmochimica Acta, 51(10), 2707-2717. https://doi.org/10.1016/0016-7037(87)90151-7
Semprebon, G. M., Rivals, F., 2010, Trends in the paleodietary habits of fossil camels from the Tertiary and Quaternary of North America: Palaeogeography, Palaeoclimatology, Palaeoecology. 295(1-2), 131-145. https://doi.org/10.1016/j.palaeo.2010.05.033
Semprebon, G. M., Sise, P. J., Coombs, M.C., 2011, Potential bark and fruit browsing as revealed by stereomicrowear analysis of the peculiar clawed herbivores known as chalicotheres (Perissodactyla, Chalicotherioidea): Journal of Mammalian Evolution, 18(1), 33-55. https://doi.org/10.1007/s10914-010-9149-3
Shearer, G., Kohl, D. H., Virginia, R., Bryan, B. A., Skeeters, J. L., Nilsen, E. T., Sharifi, M.R., Rundel, P. W., 1983, Estimates of N2-fixation from variation in the natural abundance of 15N in Sonoran Desert ecosystems: Oecologia. 56(2-3), 365-373. https://doi.org/10.1007/BF00379714
Siebe, C., Schaaf, P., Urrutia-Fucugauchi, J., 1999, Mammoth bones embedded in a late Pleistocene lahar from Popocatépetl volcano, near Tocuila, central Mexico: Geological Society of America Bulletin, 111(10), 1550-1562. https://doi.org/10.1130/0016-7606(1999)111%3C1550:MBEIAL%3E2.3.CO;2
Simkiss, K., Wilbur, K.M., 1989, Biomineralization: Cell Biology and Mineral Deposition: San Diego, CA, Academic Press, 337 p.
Smith, G. J., DeSantis, L. R., 2018, Dietary ecology of Pleistocene mammoths and mastodons as inferred from dental microwear textures: Palaeogeography, Palaeoclimatology, Palaeoecology, 492, 10-25. https://doi.org/10.1016/j.palaeo.2017.11.024
Smith, G. J., DeSantis, L. R., 2020, Extinction of North American Cuvieronius (Mammalia: Proboscidea: Gomphotheriidae) driven by dietary resource competition with sympatric mammoths and mastodons: Paleobiology, 46(1), 41-57. https://doi.org/10.1017/pab.2020.7
Solounias, N., Tariq, M., Hou, S., Danowitz, M., Harrison, 2014, A New Method of Tooth Mesowear and a Test of it on Domestic Goats: Annales Zoologici Fennici, 51(1-2), 111-118. https://doi.org/10.5735/086.051.0212
Solounias, N., Semprebon, G., 2002, Advances in the reconstruction of ungulate ecomorphology with application to early fossil equids: American Museum Novitates, 2002(3366), 1-49. https://doi.org/10.1206/0003-0082(2002)366%3C0001:AITROU%3E2.0.CO;2
Steuter, A.A., Jasch, B., Ihnen, J., Tieszen, L. L., 1990, Woodland/Grassland Boundary Changes in the Middle Niobrara Valley of Nebraska Identified by δ13C Values of Soil Origanic Matter: American Midland Naturalist, 124(2), 301-308. https://doi.org/10.2307/2426179
Stokke, S., 1999, Sex differences in feeding-patch choice in a megaherbivore: elephants in Chobe National Park, Botswana: Canadian Journal of Zoology, 77(11), 1723-1732. https://doi.org/10.1139/z99-145
Stowe, L.G., Teeri, J.A., 1978, The geographic distribution of C4 species of the Dicotyledonae in relation to climate: The American Naturalist, 112(985), 609-623. https://doi.org/10.1086/283301
Stephens, R.B., Ouimette, A.P., Hobbie, E.A., Rowe, R. J., 2022, Reevaluating trophic discrimination factors (Δδ13C and Δδ15N) for diet reconstruction: Ecological Monographs, e1525. https://doi.org/10.1002/ecm.1525
Strait, S.G., 1993, Molar microwear in extant small‐bodied faunivorous mammals: An analysis of feature density and pit frequency: American Journal of Physical Anthropology, 92(1), 63-79. https://doi.org/10.1002/ajpa.1330920106
Sukumar, R., Ramesh, R., 1992, Stable carbon isotope ratios in Asian elephant collagen: implications for dietary studies: Oecologia, 192, 536-539. https://doi.org/10.1007/BF00650328
Szabó, B., Pazonyi, P., Tóth, E., Magyari, E.K., Kiss, G.I., Rinyu, L., Futó, I., Virág, A., 2021, Pleistocene and holocene palaeoenvironmental reconstruction of the carpathian basin based on multiproxy analysis of cervid teeth: Historical Biology, 33(12), 3307-3325. https://doi.org/10.1080/08912963.2020.1863960
Taylor, L. A., Kaiser, T. M., Schwitzer, C., Müller, D. W., Codron, D., Clauss, M., Schulz, E., 2013, Detecting inter-cusp and inter-tooth wear patterns in Rhinocerotids: PloS one, 8(12), e80921. https://doi.org/10.1371/journal.pone.0080921
Teaford, M.F., Oyen, O. J., 1989, In vivo and in vitro turnover in dental microwear: American Journal of Physical Anthropology, 80(4), 447-460. https://doi.org/10.1002/ajpa.1330800405
Teaford, M. F., Walker, A., 1984, Quantitative differences in dental microwear between primate species with different diets and a comment on the presumed diet of Sivapithecus: American journal of physical anthropology, 64(2), 191-200. https://doi.org/10.1002/ajpa.1330640213
Tejada-Lara, J. V., MacFadden, B. J., Bermudez, L., Rojas, G., Salas-Gismondi, R., Flynn, J. J., 2018, Body mass predicts isotope enrichement in herbivorous mammals: Proceedings of Royal Society B, 285 (1881), 20181020. https://doi.org/10.1098/rspb.2018.1020
Theodor, J. M., Furr, R. S., 2009, High dynamic range imaging as applied to paleontological specimen photography: Palaeontologia Electronica, 12(1), 1-30.
Tipple, B.J., Meyers, S.R., Pagani, M., 2010, Carbon isotope ratio of Cenozoic CO2: A comparative evaluation of available geochemical proxies: Paleoeceanography, 25(3), PA3202. https://doi.org/10.1029/2009PA001851
Todd, N.E., Falco, N., Silva, N., Sánchez, C., 2007, Dental microwear variation in complete molars of Loxodonta africana and Elephas maximus: Quaternary International, 169, 192-202. https://doi.org/10.1016/j.quaint.2006.08.009
Tütken, T., Furrer, H., Vennemann, T. W., 2007, Stable isotope compositions of mammoth teeth from Niederweningen, Switzerland: implications for the Late Pleistocene climate, environment, and diet: Quaternary International, 164, 139-150. https://doi.org/10.1016/j.quaint.2006.09.004
Tütken, T., Kaiser, T.M., Vennemann, T., Merceron, G., 2013, Opportunistic feeding strategy for the earliest Old World hypsodont equids: evidence from stable isotope and dental wear proxies: PLoS One, 8(9), e74463. https://doi.org/10.1371/journal.pone.0074463
Uno, K. T., Fisher, D. C., Schuster, G., Wittemyer, G., Douglas-Hamilton, I., Omondi, P., Litoroh, M., Cerling, T. E., 2020, High-resolution stable isotope profiles of modern elephant (Loxodonta africana) tusk dentin and tail hair from Kenya: Implications for identifying seasonal variability in climate, ecology, and diet in ancient proboscideans: Palaeogeography, Palaeoclimatology, Palaeoecology, 559, 109962. https://doi.org/10.1016/j.palaeo.2020.109962
Uno, K. T., Fisher, D. C., Wittemyer, G., Douglas-Hamilton, I., Carpenter, N., Omondi, P., Cerling, T. E., 2020, Forward and inverse methods for extracting climate and diet information from stable isotope profiles in proboscidean molars: Quaternary International, 557, 92-109. https://doi.org/10.1016/j.quaint.2020.06.030
Uno, K. T., Bibi, F., 2022, Stable isotope paleoecology of the Baynunah Formation, in Bibi, F., Kraatz, B., Beech, M.J., Hill, A., (eds), Sands of time. Vertebrate paleobiology and paleoanthropology: Cham, Springer, 299-331. https://doi.org/10.1007/978-3-030-83883-6_18
Van der Merwe, N. J., Medina, E., 1991, The canopy effect, carbon isotope ratios and foodwebs in Amazonia: Journal of Archaeological Science, 18(3), 249-259. https://doi.org/10.1016/0305-4403(91)90064-V
Vogel, J.C., 1978, Recycling of CO2 in a forest environment: Oecologia Plantarum, 13,89-94.
Walker, A., Hoeck, H.N., Pérez, L., 1978, Microwear of mammalian teeth as an indicator of diet: Science, 201(4359), 908-910. https://doi.org/10.1126/science.684415
Way, D.A., Katul, G.G., Manzoni, S., Vico, G., 2014, Increasing water use efficiency along the C3 to C4 evolutionary pathway: a stomatal optimization perspective: Journal of Experimental Botany, 65(13), 3683-3693. https://doi.org/10.1093/jxb/eru205
Widga, C., Hodgins, G., Kolis, K., Lengyel, S., Saunders, J., Walker, J. D., & Wanamaker, A. D., 2021, Life histories and niche dynamics in late Quaternary proboscideans from midwestern North America: Quaternary Research, 100, 224-239. https://doi.org/10.1017/qua.2020.85
Wiest, L. A., Esker, D., Driese, S. G., 2016, The Waco Mammoth National Monument may represent a diminished watering-hole scenario based on preliminary evidence of post-mortem scavenging: Palaios, 31(12), 592-606. https://doi.org/10.2110/palo.2016.053
Winkler, D. E., Kubo, M. O., 2022, Inter-microscope comparability of dental microwear texture data obtained from obtained from different optical profilometers: bioRxiv, 1-19. https://doi.org/10.1101/2022.03.08.483539
Winkler, D. E., Schulz-Kornas, E., Kaiser, T. M., De Cuyper, A., Clauss, M., Tütken, T., 2019, Forage silica and water content control dental surface texture in guinea pigs and provide implications for dietary reconstruction: Proceedings of the National Academy of Sciences, 116(4), 1325-1330. https://doi.org/10.1073/pnas.1814081116
Wittemyer, G., Cerling, T. E., Douglas-Hamilton, I., 2009, Establishing chronologies from isotopic profiles in serially collected animal tissues: An example using tail hairs from African elephants: Chemical Geology, 267(1-2), 3-11. https://doi.org/10.1016/j.chemgeo.2008.08.010
Wooller, M.J., Bataille, C., Druckenmiller, P., Erickson, G.M., Groves, P., Haubenstock, N., Howe, T., Irrgehern J., Mann, D., Moon, K., Potter, B., Prohaska, T., Rasic, J., Reuther, J., Shapiro, B., Spaleta, K., Willis, A. D., 2021, Lifetime mobility of an Arctic woolly mammoth: Science, 373(6556), 806-808. https://doi.org/10.1126/science.abg1134
Zazzo, A., Bendrey, R., Vella, D., Moloney, A. P., Monahan, F. J., Schmidt, O., 2012, A refined sampling strategy for intra-tooth stable isotope analysis of mammalian enamel: Geochimica et Cosmochimica Acta, 84, 1-13. https://doi.org/10.1016/j.gca.2012.01.012
Zhang, H., Wang, Y., Janis, C. M., Goodall, R. H., Purnell, M. A., 2017, An examination of feeding ecology in Pleistocene proboscideans from southern China (Sinomastodon, Stegodon, Elephas), by means of dental microwear texture análisis: Quaternary International, 445, 60-70. https://doi.org/10.1016/j.quaint.2016.07.011
La revisión por pares es responsabilidad de la Universidad Nacional Autónoma de México.
Este es un artículo Open Access bajo la licencia CC Y-NC-SA (https://creativecommons.org/licenses/by-nc-sa/4.0/)